
by Jonathan Latham, PhD and Allison Wilson, PhD
In all the discussions of the origin of the COVID-19 pandemic, enormous scientific attention has been paid to the molecular character of the SARS-CoV-2 virus, including its novel genome sequence in comparison with its near relatives. In stark contrast, virtually no attention has been paid to the physical provenance of those nearest genetic relatives, its presumptive ancestors, which are two viral sequences named BtCoV/4991 and RaTG13.
This neglect is surprising because their provenance is more than interesting. BtCoV/4991 and RaTG13 were collected from a mineshaft in Yunnan province, China, in 2012/2013 by researchers from the lab of Zheng-li Shi at the Wuhan Institute of Virology (WIV). Very shortly before, in the spring of 2012, six miners working in the mine had contracted a mysterious illness and three of them had died (Wu et al., 2014). The specifics of this mystery disease have been virtually forgotten; however, they are described in a Chinese Master’s thesis written in 2013 by a doctor who supervised their treatment.
We arranged to have this Master’s thesis translated into English. The evidence it contains has led us to reconsider everything we thought we knew about the origins of the COVID-19 pandemic. It has also led us to theorise a plausible route by which an apparently isolated disease outbreak in a mine in 2012 led to a global pandemic in 2019.
The origin of SARS-CoV-2 that we propose below is based on the case histories of these miners and their hospital treatment. This simple theory accounts for all the key features of the novel SARS-CoV-2 virus, including ones that have puzzled virologists since the outbreak began.
The theory can account for the origin of the polybasic furin cleavage site, which is a region of the viral spike protein that makes it susceptible to cleavage by the host enzyme furin and which greatly enhances viral spread in the body. This furin site is novel to SARS-CoV-2 compared to its near relatives (Coutard, et al., 2020). The theory also explains the exceptional affinity of the virus spike protein for human receptors, which has also surprised virologists (Letko et al., 2020; Piplani et al, 2020; Wrapp et al., 2020; Walls et al., 2020). The theory further explains why the virus has barely evolved since the pandemic began, which is also a deeply puzzling aspect of a virus supposedly new to humans (Zhan et al., 2020; van Dorp et al., 2020; Chaw et al., 2020). Lastly, the theory neatly explains why SARS-CoV-2 targets the lungs, which is unusual for a coronavirus (Huang et al., 2020).
We do not propose a specifically genetically engineered or biowarfare origin for the virus but the theory does propose an essential causative role in the pandemic for scientific research carried out by the laboratory of Zheng-li Shi at the WIV; thus also explaining Wuhan as the location of the epicentre.
Why has the provenance of RaTG13 and BtCoV/4991 been ignored?
The apparent origin of the COVID-19 pandemic is the city of Wuhan in Hubei province, China. Wuhan is also home to the world’s leading research centre for bat coronaviruses. There are two virology labs in the city, both have either collected bat coronaviruses or researched them in the recent past. The Shi lab, which collected BtCoV/4991 and RaTG13, recently received grants to evaluate by experiment the potential for pandemic pathogenicity of the novel bat coronaviruses they collected from the wild.
To add to these suggestive data points, there is a long history of accidents, disease outbreaks, and even pandemics resulting from lab accidents with viruses (Furmanski, 2014; Weiss et al., 2015). For these and other reasons, summarised in our article The Case is Building that COVID-19 Had a Lab Origin, we (a virologist and a geneticist) and others have concluded that a lab outbreak is a credible thesis. Certainly, a lab origin has at least as much circumstantial evidence to support it as does any natural zoonotic origin theory (Piplani et al., 2020; Segreto and Deigin, 2020; Zhan et al., 2020).
The media, normally so enamoured of controversy, has largely declined even to debate the possibility of a laboratory escape. Many news sites have simply labelled it a conspiracy theory.
The principal reason for media dismissals of the lab origin possibility is a review paper in Nature Medicine (Andersen et al., 2020). Although by Jun 29 2020 this review had almost 700 citations it also has major scientific shortcomings. These flaws are worth understanding in their own right but they are also useful background for understanding the implications of the Master’s thesis.
Andersen et al., a critique
The question of the origin of the COVID-19 pandemic is, in outline, simple. There are two incontrovertible facts. One, the disease is caused by a human viral pathogen, SARS-CoV-2, first identified in Wuhan in December 2019 and whose RNA genome sequence is known. Second, all of its nearest known relatives come from bats. Beyond any reasonable doubt SARS-CoV-2 evolved from an ancestral bat virus. The task the Nature Medicine authors set for themselves was to establish the relative merits of each of the various possible routes (lab vs natural) by which a bat coronavirus might have jumped to humans and in the same process have acquired an unusual furin site and a spike protein having very high affinity for the human ACE2 receptor.
When Andersen et al. outline a natural zoonotic pathway they speculate extensively about how the leap might have occurred. In particular they elaborate on a proposed residence in intermediate animals, likely pangolins. For example, “The presence in pangolins of an RBD [Receptor Binding Domain] very similar to that of SARS-CoV-2 means that we can infer that this was probably in the virus that jumped to humans. This leaves the insertion of [a] polybasic cleavage site to occur during human-to-human transmission.” This viral evolution occurred in “Malayan pangolins illegally imported into Guangdong province”. Even with these speculations there are major gaps in this theory. For example, why is the virus so well adapted to humans? Why Wuhan (in Hubei province), which is 1,000 Km from Guangdong? (See map).
The authors provide no such speculations in favour of the lab accident thesis, only speculation against it:
“Finally, the generation of the predicted O-linked glycans is also unlikely to have occurred due to cell-culture passage, as such features suggest the involvement of an immune system.” (italics added).
[Passaging is the deliberate placing of live viruses into cells or organisms to which they are NOT adapted for the purpose of making them adapted, i.e. speeding up their evolution.]
It is also noteworthy that the Andersen authors set a higher hurdle for the lab thesis than the zoonotic thesis. In their account, the lab thesis is required to explain all of the evolution of SARS-CoV-2 from its presumed bat viral ancestor, whereas under their telling of the zoonotic thesis the key step of the addition of the furin site is allowed to happen in humans and is thus effectively unexplained.
A further imbalance is that key information needed to judge the merits of a lab origin theory is missing from their account. As we detailed in our previous article, in their search for SARS-like viruses with zoonotic spillover potential, researchers at the WIV have passaged live bat viruses in monkey and human cells (Wang et al., 2019). They have also performed many recombinant experiments with diverse bat coronaviruses (Ge et al., 2013; Menachery et al., 2015; Hu et al., 2017). Such experiments have generated international concern over the possible creation of potential pandemic viruses (Lipsitch, 2018). As we showed too, the Shi lab had also won a grant to extend that work to whole live animals. They planned “virus infection experiments across a range of cell cultures from different species and humanized mice” with recombinant bat coronaviruses. Yet Andersen et al did not discuss this research at all, except to say:
“Basic research involving passage of bat SARS-CoV-like coronaviruses in cell culture and/or animal models has been ongoing for many years in biosafety level 2 laboratories across the world”
This statement is fundamentally misleading about the kind of research performed at the Shi lab.
A further important oversight by the Andersen authors concerns the history of lab outbreaks of viral pathogens. They write: “there are documented instances of laboratory escapes of SARS-CoV”. This is a rather matter-of-fact allusion to the fact that since 2003 there have been six documented outbreaks of SARS from labs, not all in China, with some leading to fatalities (Furmanski, 2014).
Andersen et al might have also have noted that two major human pandemics are widely accepted to have been caused by lab outbreaks of viral pathogens, H1N1 in 1977 and Venezuelan Equine Encephalitis (summarised in Furmanski, 2014). Andersen could even have noted that literally hundreds of lab accidents with viruses have resulted in near-misses or very localised outbreaks (summarised by Lynn Klotz and Sam Husseini and also Weiss et al., 2015).
Also unmentioned were instances where a lab outbreak of an experimental or engineered virus has been plausibly theorised but remains uninvestigated. For example, the most coherent explanation for the H1N1 variant ‘swine flu’ pandemic of 2009/10 that resulted in a death toll estimated by some as high as 200,000 (Duggal et al., 2016; Simonsen et al. 2013), is that a vaccine was improperly inactivated by its maker (Gibbs et al., 2009). If so, H1N1 emerged from a lab not once but twice.
Given that human and livestock viral outbreaks have frequently come from laboratories and that many scientists have warned of probable lab escapes (Lipsitch and Galvani, 2014), and that the WIV itself has a questionable biosafety record, the Andersen paper is not an even-handed treatment of the possible origins of the COVID-19 virus.
Yet its text expresses some strong opinions: “Our analyses clearly show that SARS-CoV-2 is not a laboratory construct or a purposefully manipulated virus….It is improbable that SARS-CoV-2 emerged through laboratory manipulation of a related SARS-CoV-like coronavirus…..the genetic data irrefutably show that SARS-CoV-2 is not derived from any previously used backbone….the evidence shows that SARS-CoV2 is not a purposefully manipulated virus….we do not believe that any type of laboratory-based scenario is possible.” (Andersen et al., 2020).
It is hard not to conclude that what their paper mostly shows is that Drs. Andersen, Rambaut, Lipkin, Holmes and Garry much prefer the natural zoonotic transfer thesis. Their rhetoric is forthright but the evidence does not support that confidence.
Indeed, since the publication of Andersen et al., important new evidence has emerged that undermines their zoonotic origin theory. On May 26th the Chinese CDC ruled out the Huanan “wet” market in Wuhan as the source of the outbreak. Additionally, new research on pangolins, the favoured intermediate mammal host, suggests they are not a natural reservoir of coronaviruses (Lee et al., 2020; Chan and Zhan, 2020). Furthermore, SARS-CoV-2 was found not to replicate in bat kidney or lung cells (Rhinolophus sinicus), implying that SARS-CoV-2 is not a recently-adapted spill over Chu et al., 2020).
The Mojiang mine and the Master’s thesis
In our own search to resolve the COVID-19 origin question we chose to focus on the provenance of the coronavirus genome sequences BtCoV/4991 and RaTG13, since these are the most closely related sequences to SARS-CoV-2 (98.7% and 96.2% identical respectively). See FIG 1. (reproduced from P. Zhou et al., 2020).
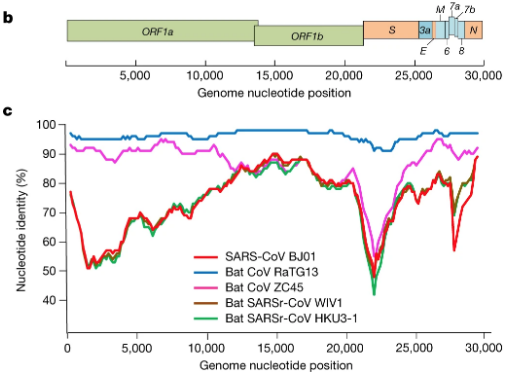
For comparison, the next closest virus to SARS-CoV-2 is RmYN02 (not shown in Fig 1.) (H. Zhou et al., 2020). RmYN02 has an overall similarity to SARS-CoV-2 of 93.2%, making its evolutionary distance from SARS-CoV-2 almost twice as great.
BtCoV/4991 was first described in 2016. It is a 370 nucleotide virus fragment collected from the Mojiang mine in 2013 by the lab of Zeng-li Shi at the WIV (Ge et al., 2016). BtCoV/4991 is 100% identical in sequence to one segment of RaTG13. RaTG13 is a complete viral genome sequence (almost 30,000 nucleotides) that was only published in 2020, after the pandemic began (P. Zhou et al., 2020).
Despite the confusion created by their different names, in a letter obtained by us Zheng-li Shi confirmed to a virology database that BtCoV/4991 and RaTG13 are both from the same bat faecal sample and the same mine. They are thus sequences from the same virus. In the discussion below we will refer primarily to RaTG13 and specify BtCoV/4991 only as necessary.
These specifics are important because it is these samples and their provenance that we believe are ultimately key to unravelling the mystery of the origins of COVID-19.
The story begins in April 2012 when six workers in that same Mojiang mine fell ill from a mystery illness while removing bat faeces. Three of the six subsequently died.
In a March 2020 interview with Scientific American Zeng-li Shi dismissed the significance of these deaths, claiming the miners died of fungal infections. Indeed, no miners or deaths are mentioned in the paper published by the Shi lab documenting the collection of RaTG13 (Ge et al., 2016).
But Shi’s assessment does not tally with any other contemporaneous accounts of the miners and their illness (Rahalkar and Bahulikar, 2020). As these authors have pointed out, Science magazine wrote up part of the incident in 2014 as A New Killer Virus in China?. Science was citing a different team of virologists who found a paramyxovirus in rats from the mine. These virologists told Science they found “no direct relationship between human infection” and their virus. This expedition was later published as the discovery of a new virus called MojV after Mojiang, the locality of the mine (Wu et al., 2014).
What this episode suggests though is that these researchers were looking for a potentially lethal virus and not a lethal fungus. Also searching the Mojiang mine for a virus at around the same time was Canping Huang, the author of a PhD thesis carried out under the supervision of George Gao, the head of the Chinese CDC.
All of this begs the question of why the Shi lab, which has no interest in fungi but a great interest in SARS-like bat coronaviruses, also searched the Mojiang mine for bat viruses on four separate occasions between August 2012 and July 2013, even though the mine is 1,000 Km from Wuhan (Ge et al., 2016). These collecting trips began while some of the miners were still hospitalised.
Fortunately, a detailed account of the miner’s diagnoses and treatments exists. It is found in a Master’s thesis written in Chinese in May 2013. Its suggestive English title is “The Analysis of 6 Patients with Severe Pneumonia Caused by Unknown viruses“.
The original English version of the abstract implicates a SARS-like coronavirus as the probable causative agent and that the mine “had a lot of bats and bats’ feces”.
The findings of the Master’s thesis
To learn more, especially about the reasonableness of this diagnosis, we arranged to have the whole Master’s thesis translated into English and are here making the translation available. To read it in full see the embedded document below (or download it here).
Analysis of Six Patients With Unknown Viruses (Text)
The six ill miners were admitted to the No. 1. School of Clinical Medicine, Kunming Medical University, in short succession in late April and early May 2012. Kunming is the capital of Yunnan province and 250 Km from Mojiang.
Of the descriptions of the miners and their treatments, which include radiographs and numerous CAT scans, several features stand out:
1) From their admission to the hospital their doctors informed the “medical office” of a potential “outburst of disease” i.e. a potential epidemic outbreak. Thus, the miners were treated for infections and not as if they had inhaled noxious gases or other toxins.
2) The symptoms (on admission) of the six miners were: a) dry cough, b) sputum, c) high fevers, especially shortly before death d) difficulty breathing, e) myalgia (sore limbs). Some patients had hiccoughs and headaches. (See Table 1).
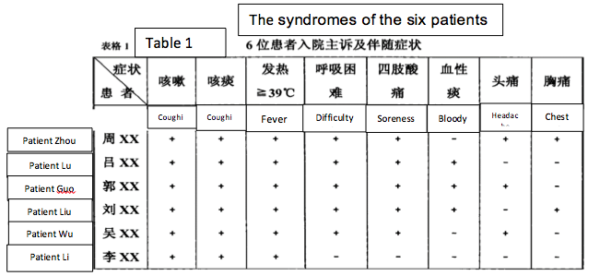
3) Clinical work established that patients 1-4 had low blood oxygen “for sure it was ARDS” (Acute Respiratory Distress Syndrome) and immune damage considered indicative of viral infection. Additionally, a tendency for thrombosis was noted in patients 2 and 4. Symptom severity and mortality were age-related (though from a sample of 6 this must be considered anecdotal).
4) Potential common and rare causes of their symptoms were tested for and mostly eliminated. For patients 3 and 4 these included tests for HIV, Cytomegalovirus, Epstein-Barr Virus (EBV), Japanese encephalitis, haemorrhagic fever, Dengue, Hepatitis B, SARS, and influenza. Of these, only patient 2 tested positive for Hepatitis and EBV.
5) Treatment of the six patients included ventilation (patients 2-4), steroids (all patients), antivirals (all except patient 5), and blood thinners (patients 2 and 4). Antibiotics and antifungal medications were administered to counter what were considered secondary (but significant) co-infections.
6) A small number of remote meetings were held with researchers at other universities. One was with Zhong Nanshan at Sun Yat-Sen University, Guangdong. Zhong is the Chinese hero of the SARS epidemic, a virologist, and arguably the most famous scientist in China.
7) Samples from the miners were later sent to the WIV in Wuhan and to Zhong Nanshan, further confirming that viral disease was strongly suspected. Some miners did test positive for coronavirus (the thesis is unclear on how many).
8) The source of infection was concluded to be Rhinolophus sinicus, a horseshoe bat and the ultimate conclusion of the thesis reads “the unknown virus lead to severe pneumonia could be: The SARS-like-CoV from the Chinese rufous horseshoe bat.” Thus the miners had a coronavirus but it apparently was not SARS itself.
The significance of the Master’s thesis
These findings of the thesis are significant in several ways.
First, in the light of the current coronavirus pandemic it is evident the miners’ symptoms very closely resemble those of COVID-19 (Huang et al, 2020; Tay et al., 2020; M. Zhou et al., 2020). Anyone presenting with them today would immediately be assumed to have COVID-19. Likewise, many of the treatments given to the miners have become standard for COVID-19 (Tay et al., 2020).
Second, the remote meeting with Zhong Nanshan is significant. It implies that the illnesses of the six miners were of high concern and, second, that a SARS-like coronavirus was considered a likely cause.
Third, the abstract, the conclusions, and the general inferences to be made from the Master’s thesis contradict Zheng-li Shi’s assertion that the miners died from a fungal infection. Fungal infection as a potential primary cause was raised but largely discarded.
Fourth, if a SARS-like coronavirus was the source of their illness the implication is that it could directly infect human cells. This would be unusual for a bat coronavirus (Ge et al., 2013). People do sometimes get ill from bat faeces but the standard explanation is histoplasmosis, a fungal infection and not a virus (McKinsey and McKinsey, 2011; Pan et al., 2013).
Fifth, the sampling by the Shi lab found that bat coronaviruses were unusually abundant in the mine (Ge at al., 2016). Among their findings were two betacoronaviruses, one of which was RaTG13 (then known as BtCoV/4991). In the coronavirus world betacoronaviruses are special in that both SARS and MERS, the most deadly of all coronaviruses, are both betacoronaviruses. Thus they are considered to have special pandemic potential, as the concluding sentence of the Shi lab publication which found RaTG13 implied: “special attention should particularly be paid to these lineages of coronaviruses” (Ge at al., 2016). In fact, the Shi and other labs have for years been predicting that bat betacoronaviruses like RaTG13 would go pandemic; so to find RaTG13 where the miners fell ill was a scenario in perfect alignment with their expectations.
The Mojiang miners passaging proposal
How does the Master’s thesis inform the search for a plausible origin of the pandemic?
In our previous article we briefly discussed how the pandemic might have been caused either by a virus collection accident, or through viral passaging, or through genetic engineering and a subsequent lab escape. The genetic engineering possibility deserves attention and is extensively assessed in an important preprint (Segreto and Deigin, 2020).
We do not definitively rule out these possibilities. Indeed it now seems that the Shi lab at the WIV did not forget about RaTG13 but were sequencing its genome in 2017 and 2018. However, we believe that the Master’s thesis indicates a much simpler explanation.
We suggest, first, that inside the miners RaTG13 (or a very similar virus) evolved into SARS-CoV-2, an unusually pathogenic coronavirus highly adapted to humans. Second, that the Shi lab used medical samples taken from the miners and sent to them by Kunming University Hospital for their research. It was this human-adapted virus, now known as SARS-CoV-2, that escaped from the WIV in 2019.
We refer to this COVID-19 origin hypothesis as the Mojiang Miners Passage (MMP) hypothesis.
Passaging is a standard virological technique for adapting viruses to new species, tissues, or cell types. It is normally done by deliberately infecting a new host species or a new host cell type with a high dose of virus. This initial viral infection would ordinarily die out because the host’s immune system vanquishes the ill-adapted virus. But, in passaging, before it does die out a sample is extracted and transferred to a new identical tissue, where viral infection restarts. Done iteratively, this technique (called “serial passaging” or just “passaging”) intensively selects for viruses adapted to the new host or cell type (Herfst et al., 2012).
At first glance RaTG13 is unlikely to have evolved into SARS-CoV-2 since RaTG13 is approximately 1,200 nucleotides (3.8%) different from SARS-CoV-2. Although RaTG13 is the most closely related virus to SARS-CoV-2, this sequence difference still represents a considerable gap. In a media statement evolutionary virologist Edward Holmes has suggested this gap represents 20-50 years of evolution and others have suggested similar figures.
We agree that ordinary rates of evolution would not allow RaTG13 to evolve into SARS-CoV-2 but we also believe that conditions inside the lungs of the miners were far from ordinary. Five major factors specific to the hospitalised miners favoured a very high rate of evolution inside them.
i) When viruses infect new species they typically undergo a period of very rapid evolution because the selection pressure on the invading pathogen is high. The phenomenon of rapid evolution in new hosts is well attested among corona- and other viruses (Makino et al., 1986; Baric et al., 1997; Dudas and Rambaut 2016; Forni et al., 2017).
ii) Judging by their clinical symptoms such as the CT scans, all the miner’s infections were primarily of the lungs. This localisation likely occurred initially because the miners were exerting themselves and therefore inhaling the disturbed bat guano deeply. As miners, they may already have had damaged lung tissues (patient 3 had suspected pneumoconiosis) and/or particulate matter was present that irritated the tissues and may have facilitated initial viral entry.
In contrast, standard coronavirus infections are confined to the throat and upper respiratory tract. They do not normally reach the lungs (Perlman and Netland, 2009). Lungs are far larger tissues by weight (kilos vs grammes) than the upper respiratory tract. There was therefore likely a much larger quantity of virus inside the miners than would be the case in an ordinary coronavirus infection.
Comparing a typical coronavirus respiratory tract infection with the extent of infected lungs in the miners from a purely mathematical point of view indicates the potential scale of this quantitative difference. The human aerodigestive tract is approximately 20cm in length and 5cm in circumference, i.e. approximately 100 cm2 in surface area. The surface area of a human lung ranges from 260,000-680,000 cm2 (Hasleton, 1972). The amount of potentially infected tissue in an average lung is therefore approximately 4500-fold greater than that available to a normal coronavirus infection. The amount of virus present in the infected miners, sufficient to hospitalise all of them and kill half of them, was thus proportionately very large.
Evolutionary change is in large part a function of the population size. The lungs of the miners, we suggest, supported a very high viral load leading to proportionately rapid viral evolution.
Furthermore, according to the Master’s thesis, the immune systems of the miners were compromised and remained so even for those discharged. This weakness on the part of the miners may also have encouraged evolution of the virus.
iii) The length of infection experienced by the miners (especially patients 2, 3 and 4) far exceeded that of an ordinary coronavirus infection. From first becoming too sick to work in the mine, patient 2 survived 57 days until he died. Patient 3 survived 120 days after stopping work. Patient 4 survived 117 days and then was discharged as cured. Each had been exposed in the mine for 14 days prior to the onset of severe symptoms; thus each presumably had nascent infections for some time before calling in sick (See Table 2 of the thesis).
In contrast, in ordinary coronavirus infections the viral infection is cleared within about ten to fourteen days after being acquired (Tay et al., 2020). Thus, unlike most sufferers from coronavirus infection, the hospitalised miners had very long-term bouts of disease characterised by a continuous high load of virus. In the cases of patients 3 and 4 their illnesses lasted over 4 months.
iv) Coronaviruses are well known to recombine at very high rates: 10% of all progeny in a cell can be recombinants (Makino et al., 1986; Banner and Lai, 1991; Dudas and Rambaut, 2016). In normal virus evolution the mutation rate and the selection pressure are the main foci of attention. But in the case of a coronavirus adapting to a new host where many mutations distributed all over the genome are required to fully adapt to the new host, the recombination rate is likely to be highly influential in determining the overall speed of adaptation by the virus population (Baric et al., 1997).
Inside the miners a large tissue was simultaneously infected by a population of poorly-adapted viruses, with each therefore under pressure to adapt. Even if the starting population of virus lacked any diversity, many individual viruses would have acquired mutations independently but only recombination would have allowed these mutations to unite in the same genome. To recombine, viruses must be present in the same cell. In such a situation the particularities of lung tissues become potentially important because the existence of airways (bronchial tubes, etc.) allows partially-adapted viruses from independent viral populations to travel to distal parts of the lung (or even the other lung) and encounter other such partially-adapted viruses and populations. This movement around the lungs would likely have resulted in what amounted to a passaging effect without the need for a researcher to infect new tissues. Indeed, in the Master’s thesis the observation is several times made that areas of the lungs of a specific patient would appear to heal even while other parts of the lungs would become infected.
v) There were also a number of unusual things about the bat coronaviruses in the mine. They were abnormally abundant but also there were many different kinds, often causing co-infections of the bats (Ge et al., 2016). Viral co-infections are often more infectious or more pathogenic (Latham and Wilson, 2007).
As the WIV researchers remarked about the bats in the mine:
“we observed a high rate of co-infection with two coronavirus species and interspecies infection with the same coronavirus species within or across bat families. These phenomena may be owing to the diversity and high density of bat populations in the same cave, facilitating coronavirus intra- and interspecies transmissions, which may result in recombination and acceleration of coronavirus evolution.” (Ge et al., 2016).
The diversity of coronaviruses in the mine suggests that the miners were similarly exposed and that their illness may potentially have begun as co-infections.
Combining these observations, we propose that the miners’ lungs offered an unprecedented opportunity for accelerated evolution of a highly bat-adapted coronavirus into a highly human-adapted coronavirus and that decades of ordinary coronavirus evolution could easily have been condensed into months. However, we acknowledge that these conditions were unique. They and their scale have no exact scientific precedent we can refer to and they would be hard to replicate in a lab; thus it is important to emphasize that our proposal is fully consistent with the underlying principles of viral evolution as understood today.
In support of the MMP theory we also know something about the samples taken from the miners. According to the Master’s thesis, samples were taken from patients for “scientific research” and blood samples (at least) were sent to the WIV.
“In the later stage we worked with Dr. Zhong Nan Shan and did some sampling. The patient* tested positive for serum IgM by the WuHan Institute of Virology. It suggested the existence of virus infection” (p62 in the section “Comprehensive Analysis”.)
(*The original does not specify the number of patients tested.)
The Master’s thesis also states its regret that no samples for research were taken from patients 1 and 2, implying that samples were taken from all the others.
We further know that, on June 27th, 2012, the doctors performed an unexplained thymectomy on patient 4. The thymus is an immune organ that can potentially be removed without greatly harming the patient and it could have contained large quantities of virus. Beyond this the Master’s thesis is unfortunately unclear on the specifics of what sampling was done, for what purpose, and where each particular sample went.
Given the interests of the Shi lab in zoonotic origins of human disease, once such a sample was sent to them, it would have been obvious and straightforward for them to investigate how a virus from bats had managed to infect these miners. Any viruses recoverable from the miners would likely have been viewed by them as a unique natural experiment in human passaging offering unprecedented and otherwise-impossible-to-obtain insights into how bat coronaviruses can adapt to humans.
The logical course of such research would be to sequence viral RNA extracted directly from unfrozen tissue or blood samples and/or to generate live infectious clones for which it would be useful (if not imperative) to amplify the virus by placing it in human cell culture. Either technique could have led to accidental infection of a lab researcher.
Our supposition as to why there was a time lag between sample collection (in 2012/2013) and the COVID-19 outbreak is that the researchers were awaiting BSL-4 lab construction and certification, which was underway in 2013 but delayed until 2018.
We propose that, when frozen samples derived from the miners were eventually opened in the Wuhan lab they were already highly adapted to humans to an extent possibly not anticipated by the researchers. One small mistake or mechanical breakdown could have led directly to the first human infection in late 2019.
Thus, one of the miners, most likely patient 3, or patient 4 (whose thymus was removed), was effectively patient zero of the COVID-19 epidemic. In this scenario, COVID-19 is not an engineered virus; but, equally, if it had not been taken to Wuhan and no further molecular research had been performed or planned for it then the virus would have died out from natural causes, rather than escaped to initiate the COVID-19 pandemic.
Evidence in favour of the MMP proposal
Our proposal is consistent with all the principal undisputed facts concerning SARS-CoV-2 and its origin. The MMP proposal has the additional benefit of reconciling many observations concerning SARS-CoV-2 that have proven difficult to reconcile with any natural zoonotic hypothesis.
For instance, using different approaches, numerous researchers have concluded that the SARS-CoV-2 spike protein has a very high affinity for the human ACE2 receptor (Walls et al., 2020; Piplani et al., 2020; Shang and Ye et al., 2020; Wrapp et al., 2020). Such exceptional affinities, ten to twenty times as great as that of the original SARS virus, do not arise at random, making it very hard to explain in any other way than for the virus to have been strongly selected in the presence of a human ACE2 receptor (Piplani et al., 2020).
In addition to this, a recent report found that the spike of RaTG13 binds the human ACE2 receptor (Shang and Ye et al., 2020). We proposed above that the virus in the mine directly infected humans lung cells. The main determinant of cell infection and species specificity of coronaviruses is initial receptor binding (Perlman and Netland, 2009). Thus RaTG13, unlike most bat coronaviruses, probably can enter and infect human cells, providing biological plausibility to the idea that the miners became infected with a coronavirus resembling RaTG13.
Moreover, the receptor binding domain (RBD) of SARS-CoV-2, which is the region of the spike that physically contacts the human ACE2 receptor, has recently been crystallised to reveal its spatial structure (Shang and Ye et al., 2020). These authors found close structural similarities between the spikes of SARS-CoV-2 and RaTG13 in how they bound the human ACE2 receptor:
“Second, as with SARS-CoV-2, bat RaTG13 RBM [a region of the RBD] contains a similar four-residue motif in the ACE2 binding ridge, supporting the notion that SARS-CoV-2 may have evolved from RaTG13 or a RaTG13-related bat coronavirus (Extended Data Table 3 and Extended Data Fig. 7). Third, the L486F, Y493Q and D501N residue changes from RaTG13 to SARS CoV-2 enhance ACE2 recognition and may have facilitated the bat-to-human transmission of SARS-CoV-2 (Extended Data Table 3 and Extended Data Fig. 7). A lysine-to-asparagine mutation at the 479 position in the SARS-CoV RBD (corresponding to the 493 position in the SARS-CoV-2 RBD) enabled SARS-CoV to infect humans. Fourth, Leu455 contributes favourably to ACE2 recognition, and it is conserved between RaTG13 and SARS CoV-2; its presence in the SARS CoV-2 RBM may be important for the bat-to-human transmission of SARS-CoV-2″ (Shang and Ye et al., 2020). (italics added)
The significance of this molecular similarity is very great. Coronaviruses have evolved a diverse set of molecular solutions to solve the problem of binding ACE2 (Perlman and Netland, 2009; Forni et al., 2017). The fact that RaTG13 and SARS CoV-2 share the same solution makes RaTG13 a highly likely direct ancestor of Sars-CoV-2.
A further widely noted feature of SARS-CoV-2 is its furin site (Coutard et al., 2020). This site is absent from RaTG13 and other closely related coronaviruses. The most closely related virus with such a site is the highly lethal MERS (which broke out in 2012). Possession of a furin site enables SARS-CoV-2 (like MERS) to infect lungs and many other body tissues (such as the gastrointestinal tract and neurons), explaining much of its lethality (Hoffman et al., 2020; Lamers et al., 2020). However, no convincing explanation for how SARS-CoV-2 acquired this site has yet been offered. Our suggestion is that it arose due to the high selection pressure which existed in the miner’s lungs and which in general worked to ensure that the virus became highly adapted to the lungs. This explanation, which encompasses how SARS-CoV-2 came to target lung tissues in general, is an important aspect of our proposal.
The implication is therefore that the furin site was not acquired by recombination with another coronavirus and simply represents convergent evolution (as suggested by Andersen et al., 2020).
An intriguing alternative possibility is that SARS-CoV-2 acquired its furin site directly from the miner’s lungs. Humans possess an epithelial sodium channel protein called ENaC-a whose furin cleavage site is identical over eight amino acids to SARS-CoV-2 (Anand et al., 2020). ENaC-a protein is present in the same airway epithelial and lung tissues infected by SARS-CoV-2. It is known from plants that positive-stranded RNA viruses recombine readily with host mRNAs (Greene and Allison, 1994; Greene and Allison, 1996; Lommel and Xiong, 1991; Borja et al., 2007). The same evidence base is not available for positive-stranded animal RNA viruses, (though see Gorbalenya, 1992) but if plant viruses are a guide then acquisition of its furin site via recombination with the mRNA which encodes ENaC-a by SARS-CoV-2 is a strong possibility.
A further feature of SARS-CoV-2 has been the very limited adaptive evolution of its genome since the pandemic began (Zhan et al., 2020; van Dorp et al., 2020; Starr et al., 2020). It is a well-established principle that viruses that jump species undergo accelerated evolutionary change in their new host (e.g. Baric et al., 1997). Thus, SARS and MERS (both coronaviruses) underwent rapid and readily detectable adaptation to their new human hosts (Forni et al., 2017; Dudas and Rambaut, 2016). Such an adaptation period has not been observed for SARS-CoV-2 even though it has now infected many more individuals than SARS or MERS did. This has even led to suggestions that the SARS-CoV-2 virus had a period of cryptic circulation in humans infections that predated the pandemic (Chaw et al., 2020). The sole mutation consistently observed to accumulate across multiple studies is a D614G substitution in the spike protein (e.g. Korber et al., 2020). The numerically largest analysis of SARS-CoV-2 genomes, however, found no evidence at all for adaptive evolution, even for D614G (van Dorp et al., 2020).
The general observation is therefore that Sars-CoV-2 has remained functionally unchanged or virtually so (except for inconsequential genetic changes) since the pandemic began. This is a very important observation. It implies that SARS-CoV-2 is highly adapted across its whole set of component proteins and not just at the spike (Zhan et al., 2020). That is to say, its evolutionary leap to humans was completed before the 2019 pandemic began.
It is hard to imagine an explanation for this high adaptiveness other than some kind of passaging in a human body (Zhan et al., 2020). Not even passaging in human cells could have achieved such an outcome.
Two examples illustrate this point. In a follow up to Shang and Ye et al., (2020), a similar group of Minnesota researchers identified a distinct strategy by which the spike (S) protein (which contains the receptor bind domain; RBD) of SARS-CoV-2 evades the human immune system (Shang and Wan et al., 2020). This strategy involves more effective hiding of its RBD, but it implies again that the spike and the RBD evolved in tandem and in the presence of the human immune system (i.e. in a human body and not in tissue culture).
The Andersen authors, in their critique of a possible engineered origin for SARS-CoV-2, also stress the need for passaging in whole humans:
“Finally, the generation of the predicted O-linked glycans is also unlikely to have occurred during cell-culture passage, as such features suggest the involvement of an immune system” (Andersen et al., 2020).
The final point that we would like to make is that the principal zoonotic origin thesis is the one proposed by Andersen et al. Apart from being poorly supported this thesis is very complex. It requires two species jumps, at least two recombination events between quite distantly related coronaviruses and the physical transfer of a pangolin (having a coronavirus infection) from outside China (Andersen et al., 2020). Even then it provides no logical explanation of the adaptedness of SARS-CoV-2 across its whole genome or why the virus emerged in Wuhan.
By contrast, our MMP proposal requires only the one species jump, which is documented in the Master’s thesis. Although we do not rule out a possible role for mixed infections in the lungs of the miners, nor the possibility of recombination between closely related variants in those lungs, nor the potential acquisition of the furin site from a host mRNA, only mutation was needed to derive SARS-CoV-2 from RaTG13. Hence our attention earlier to the figure from P. Zhou et al., 2020 showing that RaTG13 is the most closely related virus to SARS-CoV-2 over its entire length. This extended similarity is perfectly consistent with a mutational origin of SARS-CoV-2 from RaTG13.
In short, the MMP theory is a plausible and parsimonious explanation of all the key features of the COVID-19 pandemic and its origin. It accounts for the propensity of SARS-CoV-2 infections to target the lungs; the apparent preadapted nature of the virus; and its transmission from bats in Yunnan to humans in Wuhan.
Further questions
The hypothesis that SARS-CoV-2 evolved in the Mojiang miner’s lungs potentially resolves many scientific questions about the origin of the pandemic. But it raises others having to do with why this information has not come to light hitherto. The most obvious of these concern the actions of the Shi lab at the WIV.
Why did the Shi lab not acknowledge the miners’ deaths in any paper describing samples taken from the mine (Ge et al., 2016 and P. Zhou et al., 2020)? Why in the title of the Ge at al. 2016 paper did the Shi lab call it an “abandoned” mine? When they published the sequence of RaTG13 in Feb. 2020, why did the Shi lab provide a new name (RaTG13) for BtCoV/4991 when they had by then cited BtCoV/4991 twice in publications and once in a genome sequence database and when their sequences were from the same sample and 100% identical (P. Zhou et al., 2020)? If it was just a name change, why no acknowledgement of this in their 2020 paper describing RaTG13 (Bengston, 2020)? These strange and unscientific actions have obscured the origins of the closest viral relatives of SARS-CoV-2, viruses that are suspected to have caused a COVID-like illness in 2012 and which may be key to understanding not just the origin of the COVID-19 pandemic but the future behaviour of SARS-CoV-2.
These are not the only questionable actions associated with the provenance of samples from the mine. There were five scientific publications that very early in the pandemic reported whole genome sequences for SARS-CoV-2 (Chan et al., 2020; Chen et al., 2020; Wu et al., 2020; P. Zhou et al., 2020; Zhu et al., 2020). Despite three of them having experienced viral evolutionary biologists as authors (George Gao, Zheng-li Shi and Edward Holmes) only one of these (Chen et al., 2020) succeeded in identifying the most closely related viral sequence by far: BtCoV/4991 a viral sequence in the possession of the Shi lab at the WIV that differed from SARS-CoV-2 by just 5 nucleotides.
As we noted in our earlier article, the most important of the questions surrounding the origins of SARS-CoV-2 could potentially be resolved by a simple examination of the complete lab notebooks and biosafety records of relevant researchers at the WIV. Now that a credible and testable lab escape hypothesis exists this task becomes potentially much easier. This moment thus represents an opportune one to renew that call for an independent and transparent investigation of the WIV.
In requesting an investigation we are aware that no scientific institution anywhere has made a comparable request. We believe that this failure undermines public trust in a “scientific response” to the pandemic. Instead, the scientific establishment has labeled the lab escape theory a “rumor“, an “unverified theory” and a “conspiracy” when its proper name is a hypothesis. By taking this stance the scientific establishment has given the unambiguous message that scientists who take the possibility of a lab origin seriously are jeopardising their careers. Thus, while countless scientific publications on the pandemic assert in their introductions that a zoonotic origin for SARS-CoV-2 is a matter of fact or near-certainty (and Andersen et al has 860 citations as of July 14th), there is still not one published scientific paper asserting that a lab escape is even a credible hypothesis that deserves investigation.
Anyone who doubts this pressure should read the interview with Birger Sørensen in Norway’s Minerva magazine in which Sørensen discusses the “reluctance” of journals to publish his assessment that the existence of a virus that is “exceptionally well adjusted to infect humans” is “suspicious” and “cannot have evolved naturally”. The source of this reluctance, says Sørensen, is not rationality or scientific evidence. It results from conflicts of interest. This mirrors our experience. To find genuinely critical analysis of COVID-19 origin theories one has to go to Twitter, blog posts, and preprint servers. The malaise runs deep when even scientists start to complain that they don’t trust science.
We nevertheless hope that journalists will investigate some of the conflicts of interest that are keeping scientists and institutions from properly investigating the lab escape hypothesis.
References
Anand, P., Puranik, A., Aravamudan, M., Venkatakrishnan, A. J., & Soundararajan, V. (2020). SARS-CoV-2 strategically mimics proteolytic activation of human ENaC. Elife, 9, e58603.
Andersen, K. G., Rambaut, A., Lipkin, W. I., Holmes, E. C., & Garry, R. F. (2020). The proximal origin of SARS-CoV-2. Nature medicine, 26(4), 450-452.
Banner, L. R., & Mc Lai, M. (1991). Random nature of coronavirus RNA recombination in the absence of selection pressure. Virology, 185(1), 441-445.
Baric, R. S., Yount, B., Hensley, L., Peel, S. A., & Chen, W. A. N. (1997). Episodic evolution mediates interspecies transfer of a murine coronavirus. Journal of virology, 71(3), 1946-1955.
Becker, Y. (2000). Evolution of viruses by acquisition of cellular RNA or DNA nucleotide sequences and genes: an introduction. Virus Genes, 21(1-2), 7-12.
Bengston, D. (2020). All journal articles evaluating the origin or epidemiology of SARS-CoV-2 that utilize the RaTG13 bat strain genomics are potentially flawed and should be retracted. OSF Preprint: https://osf.io/wy89d
Borja, M., Rubio, T., Scholthof, H. B., & Jackson, A. O. (1999). Restoration of wild-type virus by double recombination of tombusvirus mutants with a host transgene. Molecular Plant-Microbe Interactions, 12(2), 153-162.
Chan, J. F. W., Kok, K. H., Zhu, Z., Chu, H., To, K. K. W., Yuan, S., & Yuen, K. Y. (2020). Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerging microbes & infections, 9(1), 221-236.
Chaw, S. M., Tai, J. H., Chen, S. L., Hsieh, C. H., Chang, S. Y., Yeh, S. H., … & Wang, H. Y. (2020). The origin and underlying driving forces of the SARS-CoV-2 outbreak. Journal of biomedical science, 27(1), 1-12.
Chen, L., Liu, W., Zhang, Q., Xu, K., Ye, G., Wu, W., … & Mei, Y. (2020). RNA based mNGS approach identifies a novel human coronavirus from two individual pneumonia cases in 2019 Wuhan outbreak. Emerging microbes & infections, 9(1), 313-319.
Coutard, B., Valle, C., de Lamballerie, X., Canard, B., Seidah, N. G., & Decroly, E. (2020). The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral research, 176, 104742.
Dudas, G., & Rambaut, A. (2016). MERS-CoV recombination: implications about the reservoir and potential for adaptation. Virus evolution, 2(1).
Duggal, A., Pinto, R., Rubenfeld, G., & Fowler, R. A. (2016). Global variability in reported mortality for critical illness during the 2009-10 influenza A (H1N1) pandemic: a systematic review and meta-regression to guide reporting of outcomes during disease outbreaks. PloS one, 11(5), e0155044.
Forni, D., Cagliani, R., Clerici, M., & Sironi, M. (2017). Molecular evolution of human coronavirus genomes. Trends in microbiology, 25(1), 35-48.
Furmanski, M. (2014). Laboratory Escapes and “Self-fulfilling prophecy” Epidemics. Report: Center for Arms Control and Nonproliferation. PDF available at: https://armscontrolcenter.org/wp-content/uploads/2016/02/Escaped-Viruses-final-2-17-14-copy.pdf
Ge, X. Y., Li, J. L., Yang, X. L., Chmura, A. A., Zhu, G., Epstein, J. H., … & Zhang, Y. J. (2013). Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature, 503(7477), 535-538.
Ge, X. Y., Wang, N., Zhang, W., Hu, B., Li, B., Zhang, Y. Z., … & Wang, B. (2016). Coexistence of multiple coronaviruses in several bat colonies in an abandoned mineshaft. Virologica Sinica, 31(1), 31-40.
Gibbs, A. J., Armstrong, J. S., & Downie, J. C. (2009). From where did the 2009’swine-origin’influenza A virus (H1N1) emerge?. Virology journal, 6(1), 207.
Gorbalenya, A. E. (1992). Host-related sequences in RNA viral genomes. In Seminars in Virology (Vol. 3, pp. 359-359). HARCOURT BRACE JOVANOVICH.
Greene, A. E., & Allison, R. F. (1994). Recombination between viral RNA and transgenic plant transcripts. Science, 263(5152), 1423-1425.
Greene, A. E., & Allison, R. F. (1996). Deletions in the 3′ untranslated region of cowpea chlorotic mottle virus transgene reduce recovery of recombinant viruses in transgenic plants. Virology, 225(1), 231-234.
Hasleton, P. S. (1972). The internal surface area of the adult human lung. Journal of anatomy, 112(Pt 3), 391.
Herfst, S., Schrauwen, E. J., Linster, M., Chutinimitkul, S., de Wit, E., Munster, V. J., … & Rimmelzwaan, G. F. (2012). Airborne transmission of influenza A/H5N1 virus between ferrets. science, 336(6088), 1534-1541.
Hoffmann, M., Kleine-Weber, H., & Pöhlmann, S. (2020). A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells. Molecular Cell.
Hu, B., Zeng, L. P., Yang, X. L., Ge, X. Y., Zhang, W., Li, B., … & Luo, D. S. (2017). Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS pathogens, 13(11), e1006698.
Huang, Canping (2016) Novel Virus Discovery in Bat and the Exploration of Receptor of Bat Coronavirus HKU9. PhD thesis (Original in Chinese). National Institute for Viral Disease Control and Prevention, Chinese Center for Disease Control and Prevention, June 2016. Accessed on July 15, 2020: https://eng.oversea.cnki.net/kcms/detail/detail.aspx?dbcode=CDFD&QueryID=4&CurRec=1&dbname=CDFDLAST2018&filename=1017118517.nh
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., … & Cheng, Z. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The lancet, 395(10223), 497-506.
Korber, B., Fischer, W., Gnanakaran, S. G., Yoon, H., Theiler, J., Abfalterer, W., … & Partridge, D. G. (2020). Spike mutation pipeline reveals the emergence of a more transmissible form of SARS-CoV-2. bioRxiv.
Lamers, M. M., Beumer, J., van der Vaart, J., Knoops, K., Puschhof, J., Breugem, T. I., … & van Donselaar, E. (2020). SARS-CoV-2 productively infects human gut enterocytes. Science.
Latham, J. R., & Wilson, A. K. (2008). Transcomplementation and synergism in plants: implications for viral transgenes?. Molecular Plant Pathology, 9(1), 85-103.
Latinne, A., Hu, B., Olival, K. J., Zhu, G., Zhang, L., Li, H., … & Li, B. (2020). Origin and cross-species transmission of bat coronaviruses in China. bioRxiv.
Lee, J., Hughes, T., Lee, M. H., Field, H., Rovie-Ryan, J. J., Sitam, F. T., … & Lasimbang, H. (2020). No evidence of coronaviruses or other potentially zoonotic viruses in Sunda pangolins (Manis javanica) entering the wildlife trade via Malaysia. bioRxiv.
Letko, M., Marzi, A., & Munster, V. (2020). Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nature microbiology, 5(4), 562-569.
Li, W., Shi, Z., Yu, M., Ren, W., Smith, C., Epstein, J. H., … & Zhang, J. (2005). Bats are natural reservoirs of SARS-like coronaviruses. Science, 310(5748), 676-679.
Lipsitch, M. (2018). Why Do Exceptionally Dangerous Gain-of-Function Experiments in Influenza?. In Influenza Virus (pp. 589-608). Humana Press, New York, NY.
Lipsitch, M., & Galvani, A. P. (2014). Ethical alternatives to experiments with novel potential pandemic pathogens. PLoS Med, 11(5), e1001646.
Lommel, A., & Xiong, Z. (1991). Reconstitution of a functional red clover necrotic mosaic virus by recombinational rescue of the cell-to-cell movement gene expressed in a transgenic plant. Journal of Cellular Biochemistry A, 15, 151.
Lu, R., Zhao, X., Li, J., Niu, P., Yang, B., Wu, H., … & Bi, Y. (2020). Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The Lancet, 395(10224), 565-574.
Makino, S. H. I. N. J. I., Keck, J. G., Stohlman, S. A., & Lai, M. M. (1986). High-frequency RNA recombination of murine coronaviruses. Journal of Virology, 57(3), 729-737.
McKinsey, D. S., & McKinsey, J. P. (2011, December). Pulmonary histoplasmosis. In Seminars in respiratory and critical care medicine (Vol. 32, No. 06, pp. 735-744). © Thieme Medical Publishers.
Menachery, V. D., Yount, B. L., Debbink, K., Agnihothram, S., Gralinski, L. E., Plante, J. A., … & Randell, S. H. (2015). A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nature medicine, 21(12), 1508-1513.
Pan, B., Chen, M., Pan, W., & Liao, W. (2013). Histoplasmosis: a new endemic fungal infection in China? Review and analysis of cases. Mycoses, 56(3), 212-221.
Perlman, S., & Netland, J. (2009). Coronaviruses post-SARS: update on replication and pathogenesis. Nature reviews microbiology, 7(6), 439-450.
Piplani, S., Singh, P. K., Winkler, D. A., & Petrovsky, N. (2020). In silico comparison of spike protein-ACE2 binding affinities across species; significance for the possible origin of the SARS-CoV-2 virus. arXiv preprint arXiv:2005.06199.
Rahalkar, M.C.; Bahulikar, R.A. Understanding the Origin of ‘BatCoVRaTG13’, a Virus Closest to SARS-CoV-2. Preprints 2020, 2020050322 https://www.preprints.org/manuscript/202005.0322/v2
Segreto, R., & Deigin, Y. Is considering a genetic-manipulation origin for SARS-CoV-2 a conspiracy theory that must be censored?. https://www.researchgate.net/profile/Rossana_Segreto/publication/340924249_Is_considering_a_genetic-manipulation_origin_for_SARS-CoV-2_a_conspiracy_theory_that_must_be_censored/links/5ed7c17992851c9c5e74f7dc/Is-considering-a-genetic-manipulation-origin-f
Shang, J., Wan, Y., Luo, C., Ye, G., Geng, Q., Auerbach, A., & Li, F. (2020). Cell entry mechanisms of SARS-CoV-2. Proceedings of the National Academy of Sciences, 117(21), 11727-11734.
Shang, J., Ye, G., Shi, K., Wan, Y., Luo, C., Aihara, H., … & Li, F. (2020). Structural basis of receptor recognition by SARS-CoV-2. Nature, 581(7807), 221-224.
Simonsen, L., Spreeuwenberg, P., Lustig, R., Taylor, R. J., Fleming, D. M., Kroneman, M., … & Paget, W. J. (2013). Global mortality estimates for the 2009 Influenza Pandemic from the GLaMOR project: a modeling study. PLoS Med, 10(11), e1001558.
Starr, T. N., Greaney, A. J., Hilton, S. K., Crawford, K. H., Navarro, M. J., Bowen, J. E., … & Bloom, J. D. (2020). Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. BioRxiv.
Tay, M. Z., Poh, C. M., Rénia, L., MacAry, P. A., & Ng, L. F. (2020). The trinity of COVID-19: immunity, inflammation and intervention. Nature Reviews Immunology, 1-12.
van Dorp, L., Richard, D., Tan, C. C., Shaw, L. P., Acman, M., & Balloux, F. (2020). No evidence for increased transmissibility from recurrent mutations in SARS-CoV-2. bioRxiv. doi: https://doi.org/10.1101/2020.05.21.108506
https://www.biorxiv.org/content/10.1101/2020.05.21.108506v1.abstract
Wadman, M., Couzin-Frankel, J., Kaiser, J., & Matacic, C. (2020). A rampage through the body. Science, 368(6489), 356-360.
Walls, A. C., Park, Y. J., Tortorici, M. A., Wall, A., McGuire, A. T., & Veesler, D. (2020). Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell, 180, 281-292.
Wang, N., Luo, C., Liu, H., Yang, X., Hu, B., Zhang, W., … & Peng, C. (2019). Characterization of a new member of alphacoronavirus with unique genomic features in rhinolophus bats. Viruses, 11(4), 379.
Weiss, S., Yitzhaki, S., & Shapira, S. C. (2015). Lessons to be Learned from Recent Biosafety Incidents in the United States. The Israel Medical Association Journal: IMAJ, 17(5), 269-273.
Wrapp, D., Wang, N., Corbett, K. S., Goldsmith, J. A., Hsieh, C. L., Abiona, O., … & McLellan, J. S. (2020). Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science, 367(6483), 1260-1263.
Wu, Z., Yang, L., Yang, F., Ren, X., Jiang, J., Dong, J., … & Jin, Q. (2014). Novel henipa-like virus, Mojiang paramyxovirus, in rats, China, 2012. Emerging infectious diseases, 20(6), 1064.
Wu, F., Zhao, S., Yu, B., Chen, Y. M., Wang, W., Song, Z. G., … & Yuan, M. L. (2020). A new coronavirus associated with human respiratory disease in China. Nature, 579(7798), 265-269.
Xu, Li (2013) The analysis of 6 patients with severe pneumonia caused by unknown viruses. MSc thesis (Original in Chinese). Emergency Department and EICU, The 1st Affiliated Hospital of Kunming Medical University, Kunming. Accessed on July, 15, 2020: http://eng.oversea.cnki.net/Kcms/detail/detail.aspx?filename=1013327523.nh&dbcode=CMFD&dbname=CMF D2014
Zhan, S. H., Deverman, B. E., & Chan, Y. A. (2020). SARS-CoV-2 is well adapted for humans. What does this mean for re-emergence?. bioRxiv. doi: https://doi.org/10.1101/2020.05.01.073262
Zhang, L., Jackson, C. B., Mou, H., Ojha, A., Rangarajan, E. S., Izard, T., … & Choe, H. (2020). The D614G mutation in the SARS-CoV-2 spike protein reduces S1 shedding and increases infectivity. bioRxiv
Zhang, Y. Z., & Holmes, E. C. (2020). A genomic perspective on the origin and emergence of SARS-CoV-2. Cell.
Zhou, H., Chen, X., Hu, T., Li, J., Song, H., Liu, Y., … & Shi, W. (2020). A novel bat coronavirus reveals natural insertions at the S1/S2 cleavage site of the Spike protein and a possible recombinant origin of HCoV-19. bioRxiv.
Zhou, P., Yang, X. L., Wang, X. G., Hu, B., Zhang, L., Zhang, W., … & Chen, H. D. (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. nature, 579(7798), 270-273.
Zhou, M., Zhang, X., & Qu, J. (2020). Coronavirus disease 2019 (COVID-19): a clinical update. Frontiers of medicine, 1-10.
Zhu, N., Zhang, D., Wang, W., Li, X., Yang, B., Song, J., … & Niu, P. (2020). A novel coronavirus from patients with pneumonia in China, 2019. New England Journal of Medicine.
Editor’s note 1. Several important updates on the theory are available
2) https://www.independentsciencenews.org/commentaries/phylogeographic-mapping-of-newly-discovered-coronaviruses-pinpoints-direct-progenitor-of-sars-cov-2-as-originating-from-mojiang/
3) Talk to the BMJ on virus hyperevolution in long-term COVID patients https://www.youtube.com/watch?v=eLSv4Iwk_jM&t=2988s
Editor’s note 2. We welcome comments and information about the subject of this article. However, please note that the “reply” function in the comments section is not working for people without high level access to the website. There are two possible solutions for readers wanting to reply to specific comments:
1) Enter your comment but name the commenter you are responding to (if necessary with the date of their comment). Or,
2) Mail your comment to the editor: [email protected] and they will post it as a reply. Please be sure to say who/what you are replying to.
If this article was useful to you please consider sharing it with your networks.

Very convincing arguments for the provenance, thank you for researching and sharing.
If lungs are such great places for the virus to better adapt to human hosts what chance do we have against it now that so many millions of lungs are infected?
What is the source of the translation? How did you become aware of the Master’s Thesis?
We hired a Chinese speaker to translate the Chinese Master’s thesis for us quickly, but clearly enough so we could understand the diagnosis and treatment of the miners’ illnesses. The translator was not a doctor, so a doctor kindly checked the English translation to improve clarity/accuracy of the medical terms. We became aware of the Chinese Master’s thesis and later the Chinese PhD thesis (that both reported on the mine incident) after reading the two extremely informative preprints — Rahalkar and Bahulikar 2020 and Segreto and Deigin 2020 — both referenced in the our article. Both are well worth reading.
Your assumption “…BtCoV/4991 and RaTG13 are both from the same bat faecal sample and the same mine. They are thus sequences from the same virus” is suspect at best. BtCoV/4991 only has 373 base pairs. How can you be sure that there were no significant mutations between the unpublished BtCov/4991 portion and RaTG13?
The sequences of both are publicly available so it’s not an assumption. They are identical.
excerpt
“As we noted in our earlier article, the most important of the questions surrounding the origins of SARS-CoV-2 could potentially be resolved by a simple examination of the complete lab notebooks and biosafety records of relevant researchers at the WIV.”
Why haven’t these people reconciled the issue with the Bat Woman?
https://www.scientificamerican.com/article/how-chinas-bat-woman-hunted-down-viruses-from-sars-to-the-new-coronavirus1/
excerpt
“Back in Wuhan, where the lockdown was finally lifted on April 8, China’s bat woman is not in a celebratory mood. She is distressed because stories from the Internet and major media have repeated a tenuous suggestion that SARS-CoV-2 accidentally leaked from her lab—despite the fact that its genetic sequence does not match any her lab had previously studied. Other scientists are quick to dismiss the allegation. “Shi leads a world-class lab of the highest standards,” Daszak says.”
Either she’s lying or the virus is natural.
Ah, sorry I didn’t realize they had published the entire BtCoV/4991 sequence.
Your question is not silly though. Sequencing can have errors, for example. But even if it was a tiny bit different most people would still say it was the same virus on the grounds that its not different enough to be a new strain or species.
We hired a Chinese speaker to translate the Chinese Master’s thesis for us quickly, but clearly enough so we could understand the diagnosis and treatment of the miners’ illnesses. The translator was not a doctor, so a doctor kindly checked the English translation to improve clarity/accuracy of the medical terms. We became aware of the Chinese Master’s thesis and later the Chinese PhD thesis (that both reported on the mine incident) after reading the two extremely informative preprints — Rahalkar and Bahulikar 2020 and Segreto and Deigin 2020 — both referenced in the our article. Both are well worth reading.
Preprint by Xiaoxu Sean Lin and Shihong Chen “Major concerns on the identification of bat coronavirus strain RaTG13 and quality of related Nature paper” doi:10.20944/preprints202006.0044.v1
Excellent question. We agree that questions can be asked about RaTG13. The main one for us is that it is not an infectious clone. I.e. it has not fulfilled Koch’s postulates of being able to infect a bat, cause disease, and fulfill its lifecycle. Therefore it may be a mutant or defective in some way.
” Rahalkar and Bahulikar 2020 and Segreto and Deigin 2020″
Have you contacted the authors?
(Looks like the comments I left yesterday didn’t make it for some reason–perhaps you misunderstood my enthusiasm. I’ll rephrase, be less “colorful,” and try again.)
I think this is an amazingly well-researched and well-thought out coherent piece! Thank you!
(1) About this whole miner saga, this is the first time I’ve heard about the existence of the human tissue and blood samples! If proved true, I think this is incredibly significant!
Do you have witnesses and/or paper trails that could testify to the existence of these samples?
If true, the degree of deception involved in hiding this fact seems mind-boggling to me and this culpability should be highlighted.
In all their public utterance, Shi and company have only cryptically mentioned a tiny sample of bat poop. And have indirectly claimed that the sample is in fact used up, so all traces of anything are gone!
At no place was there ever mention of a big stash of human tissue samples that are, at least according to this piece, likely heavily laden with the virus(es) in question!
The whole legend surrounding the “batwoman” mystique would have been almost an entire fraud, since the most important information was in the abundant and readily available human samples that were shipped to her labs with industrial efficiency, not in the tiny, potentially heavily contaminated poop bits that needed to be painstakingly coaxed out of stinky dark caves!
Exactly what has happened to these samples, I hope, will at some point, become a focal point of inquiry!
Exactly which strain(s) of the virus are in these samples is an incredibly intriguing question. To what extent, if at all, are they different from today’s SARS-CoV-2 or its bat ancestors? Does it include the all-important furin site?
If the samples were destroyed, who did it? Why?
(2) Just WHY?!
The all-important question of why, why did Shi and company hide this entire saga and the existence of the samples, for all these years, the all-important question of *motive*.
I understand it’s kind of beyond the scope of this scientific piece so it was probably prudent for the authors not to go there. But it’s an obvious question at least a reader should ask!
BSL4 is a means; it’s not an end. Waiting for BSL4 is waiting for a means to do what?
The work required for publication, if indeed deemed to be too risky to be carried out in China at the time, could easily be done at any number of overseas facilities, with the aid of Shi’s many international collaborators. Even more dangerous viruses had been studied safely and surely the means existed.
Shi’s entire career is built on the premise that another bat virus after Sars could one day jump from bats to human. She’s been jumping up and down telling the whole world about it. The miner saga is the holy grail! To sit on this for half a decade and keeping quiet about it is not in her character. Sitting on such a dangerous time bomb and being quiet about it is not the best way to warn the world about its impending doom.
And after the miner incident in 2013, she continued splicing experiments, creating worse and worse hybrids. The justification for the splicing experiments was that, since they couldn’t find actual natural cases of bat viruses jumping into humans, they could benefit from artificially creating them so they could study them. The miner incident showed that such jumps did in fact occur in nature and it was indeed as bad as it could get and they had all the samples they needed. That should have removed the excuses for continued splicing experiments.
In summary, I fail to see what legimate work they could possibly have intended that justified waiting for so long and so patiently to perform!
(3) “RaTG13” is a term whose meaning is no longer clear.
It could have been from the bat poop, which, critically, could be very different from what’s in the miner samples, especially in light of the main thesis of this piece, that there had been significant adaptation inside the miner lungs.
It could have come from the miners’ samples.
It could be a hodge-podge of several things in the bat poop.
And, of course, it could even have been faked!
Considering a strong condemnation has been brought against the conduct of Shi and company, the sole source of the dubious information concerning RaTG13, we can no longer confidently invest any meaning or trust in even the existence of RaTG13, let alone the finer details of its essential characteristics.
Reading this piece on its first day of publication has been a privilege and an eye-opener! Thank you!
One question that immediately comes to mind is that if these patients in 2012 had CoViD-19 then why did this not lead at least to wider local outbreaks – for example in the places where they were treated or amongst their families or the people who transported them to doctors and hospitals. Is there any information about that available?
Brilliant and very intriguing analysis. Came here through https://swprs.org/covid-19-virus-origin-the-mojiang-miners-passage-hypothesis/
They link to another worrying article: https://armswatch.com/project-g-2101-pentagon-biolab-discovered-mers-and-sars-like-coronaviruses-in-bats/
So “Eco Health Alliance” was working with both the WIV and the Pentagon.
To answer Philip Ward: Good question. Our best guess is that it took a while for virus to evolve to be highly transmissible from human to human — this most likely happened later, after the miner(s) were in hospital. Also, once the miners were in the hospital the thesis is clear they were treated as if they had infectious diseases — perhaps even SARS or other very deadly diseases. Thus we expect the hospital staff were taking every precaution. There are other possibilities — the others were infected but had no symptoms or that infections were not made public or that we just have not encountered the data yet.
Being a scientist but not a research scientist, it seems from the evidence so far that the miners were infected by an undeveloped strain of Covid-19. Even assuming that RaTG13 is natural and its postulated genome is correct and not a hypothesised extrapolation from the 370 nucleotide sample from BAT4991 (neither of which appears to have any surviving examples despite being listed as being used for research numerous times in 2017 and 2018 in the Chinese research database) there is ample scientific evidence in related research quoted in the above article that it would have taken between 20 -50 years for the RaTG13 genome to have mutated naturally to the C0vid-19 genome.
However, in 2013 previous coronoviruses had not had the human to human transmission of Covid-19 and you would not have expected doctors, nurses, ancilliary staff, let alone simple porters, stretcher bearers and good samaritans to have taken the infection precautions we now take automatically – and yet none appears to have been infected. This means at that time human to human transmission was not possible – because that part of the virus’s genome had not come to fruition.
From 2013 to 2019 various Gain of Function processes were actioned on Bat samples in the Wuhan lab to make the samples more infectious to humans in order, as the researchers are quick to explain, to enable vaccines or preventative infectious measures to become readily available. In seven years what sort of experiments could have been done to arrive at Covid-19 – look at what has been done just in the last 8 months to counter the infection. See Timothy Stout’s comments for the range of options.
What I would like to know is how the infections in Wuhan, mostly type B virus with some A and the infections in Guangdong, all type A tie up and the timeframes involved. Did researchers from Wuhan travel to Guangdong and infect locals and did the virus then mutate from A to B as A was less infectious in the Chinese hosts (possibly from previous coronavirus infections) and therefore the virus had to change. Much to be discussed.
Thank you so much for the effort to find and translate a very interesting thesis, and developing a new hypothesis.
I had questions on the time related epidemiology, and also on the question raised by Philip above. I have questions about the ability to extract full sequences some months after the infection event, based on government reporting from my home state.
The article read as though all miners passaged RaTG13 for the same outcome. One would think there’d be an infection precedent. On the hypothesis that the miners suffered from Sars-Cov-2, I would think that one would’ve passaged it first.
[The typcial reported course of the virus… There is average of 6 days from infection to mild symptoms (2-26?). Another week to symptoms becoming “serious” enough to require hospitalisation.]
Patients were admitted 26/4 (1), 25/4 (2), 27/4 (3), 26/4 (4), 2/5 (5), 26/4 (6). Patients (1)-(4) began at the mine on 2/4 and worked 2 weeks. Patients (5) and (6) began on 22/4, supposedly after (1)-(4) had left. I would ask questions about residence and contact. It’s possible the one of the first miners passaged the most successful virus, and working an enclosed space passed it to the others within a few days. Certainly patient (6) proceeded more rapidly to hospital (patient (5) quite quickly), so it would suggest they already had the virus. I’d be asking questions about where they lived – local or fly in/fly out? Do they live in a town with others? Is there any local report of doctors/hospital staff becoming infected? I suppose China had been through sars and were highly precautionary – this didn’t apply in Wuhan where there were many infections in medical staff, but perhaps this was a consequence of an avalanche of cases and low ppe availability. It’s said that 80% of cases pass the virus on to no-one and that 20% are the super-spreaders. Perhaps the 6 weren’t the spreading types?
If this was the Sars-Cov-2 virus, one would be looking for the greater prevalence of low symptom sufferers, understanding that the miners were a relatively unwell population.
Regarding the ability to extract full sequences some months after the infection event. Here in Victoria Australia there is a general view that people are non-infectious 10-14 days after first symptoms. They explain subsequent positive tests as being related to viral particles, not whole live viruses. They may not be right, or it may relate specifically to the tests they are using, but they lead me to ask questions about the extraction of full viral material from the thymus so long after the infection events.
I hope I have understood your question correctly but our contention is that, while normal coronavirus passes through a person quickly, these miners had the disease continuously for months. They did not manage to clear it and there was a kind of stalemete between the virus and the immune system that took a long time to resolve, either in death or recovery. We don’t rule out that they may have passed it to each other. Jonathan
Presumably the escapee came from one miner, specifically. They may have worked with tissues from each and compared them, but they are effectively independent viruses after months in one lung. Re rescuing virus sequences and genomes, it can be done: https://www.mdpi.com/1999-4915/11/4/379/htm
The fact that the miners were sick with Covid-19-like symptoms doesn’t preclude lab development to enhance human affinity. What they presented with may have been the symptoms of BtCoV/4991 and RaTG13. Although these coronaviruses may not have strong human affinity it may have been sufficient in damaged lungs. This could explain the lack of a public epidemic/health care worker report of illness. Are there any reports of anyone having been infected with BtCoV/4991 and RaTG13?
There are not. In the case of 4991 because it is not a complete genome.
Thank you for a very interesting article, and for providing a link to my interview with Birger Sørensen in Minerva.
I have recently written two more articles on the works of Sørensen, you might find them interesting.
This one is about the struggle to publish his article:
https://www.minervanett.no/angus-dalgleish-birger-sorensen-coronavirus/the-fight-for-a-controversial-article/362519
The second one contains an upload of his full thesis:
https://www.minervanett.no/angus-dalgleish-birger-sorensen-coronavirus/the-evidence-which-suggests-that-this-is-no-naturally-evolved-virus/362529
Lance thank you for the link to Arms Watch and the excellent article connecting EcoHealth Alliance (which has links to the Gates Foundation) to the US Military. I hope many people will read this article and the Arms Watch website http://armswatch.com/. EcoHealth gets the vast majority of its money from the government but it also has corporate funding and foundation funding which it would be useful to know more about.
@Jonathan and Allison: Prof Balloux notes that “4 out of 7 known hCoVs have furin cleavage site”, including “common cold” viruses OC43 and HKU1. Is this correct or is there a difference to SARS2 and MERS that makes them special or requires “special explanation”?
https://twitter.com/BallouxFrancois/status/1283962325746233344
There may exist other reports on the Mojiang Mine Outbreak, or reports of people other than the miners being infected. However, the only reports we are aware of are 1. The Master’s thesis by Xu,Li (2013) 2. The Huang, Canping (2016) PhD thesis(chapter 3). 3. The Science news item “A New Killer Virus in China?” (this report only mentions the three miners who died) and 4. the short scientific report by Wu et al. 2014. The originals and/or translations of these documents are all linked to in the text and/or cited in the references of our article. We would like to hear about any other documented reports on this outbreak that people can find.
alright this seems to answers it: no other CoV has ACE2/FCS combination:
https://twitter.com/flavinkins/status/1284338683676516352
https://twitter.com/flavinkins/status/1284337537473904640
So FCS not unique in general but in hCov group 2B.
This excellent piece has at least three key ideas or revelations. (1) The mine saga and human samples, a bombshell! (2) Significant and rapid adaptation by the virus in miner lungs. (3) ALL, not just some, features of the virus (including the all-important furin site) were acquired solely in miner lungs. Some may deem (2) and/or (3) controversial.
But one of the greatest contributions this piece could make is to alert people to a line of possible forensic inquiry that has a clear origin and narrative that should be pursued. We now have a solid lead, starting with the mine, to the hospital, to WIV. A lot of people were involved in this chain of events. There should be witnesses, paper trails, and medical samples. That these leads need to be pursued is entirely independent of whether one believes Idea (2) or Idea (3). The corrupt forces will likely want to confuse the matter. They would point to the potential controversies of Idea (2) or Idea (3) to discredit the entire piece and therefore obstruct an inquiry that starts with just Idea (1). People need to be aware of that and call that out if/when it happens.
And for people who believe an entirely man-made narrative, please be mindful that Idea (1) and Idea (2) are still compatible with later human manipulation. You wouldn’t want to go out of your way to discredit this piece just because you don’t believe in Idea (3). Unlike the disgraceful Andersen piece, this piece is your friend, not your enemy. The mine origin story could be one of the very few, if not the only, credible forensic leads that could be the starting point of an inquiry. If an investigation that starts with the mine ultimately leads to uncovering crimes that more closely resemble your pet theory than Idea (3), I believe the authors of this piece would have no problem, neither should you. We should be objective and critical about scientific details, but we should be united in our call for an inquiry that follows the trail leading from the mine, to the hospital, to WIV, and hopefully, to the ultimate truth!
How about the presence of a snippet of HIV in SARS-CoV-2? Does that exist? Does it point to some manipulation in the WIV lab?
We have not explored that possibility in any detail. The Shi lab did use HIV though in Wang et al. 2019: https://www.mdpi.com/1999-4915/11/4/379/htm
The story is convincing in detail, but fails to support the conclusions. The story says that Covid19 evolved naturally in miners lungs in a hospital, then they concluded that it originated from a lab. According to the detail, the lab merely stored the virus and did no significant originating work on it before it escaped. this does NOT make the lab the origin of the virus, merely a multi-year storage point during which time it seems to have been eradicated elsewhere.
SARS-CoV-2, viruses that are suspected to have caused a COVID-like illness in 2012 and which may be key to understanding not just the origin of the COVID-19 pandemic but the future behaviour of SARS-CoV-2. Well said as Dr Yan limeng in the Fox News interview warn us we need find the truth.
PS Apologies that the “REPLY” function seems not to be working. Attempting to fix this.
A kind reader (John) has made this available as a Talking Paper to download and listen to. Hear his nice English accent at:
https://soundcloud.com/talkingpapers/proposed-origin-for-sarscov2
Also available on youtube:
https://youtu.be/l3gQdkWn9u0
(link updated)
Would it be possible to publish the paper on mainstream science journals? If not, it’s unlikely to lead to an unbiased investigation, and the article will be ignored, as Dr. Sorensen’s.
The conundrum of peer review is that it can take a long time and may not be fair. One of the things we have noticed in submitting ‘challenging’ papers is that editors (and/or reviewers) often take even longer than normal. It can take six months to get a “No” and that No is sometimes based on a trivial critique. We therefore don’t cite Sorensen just because he says it. We have personal experience. This paper of ours, for example ended up never being published even though no one (including peer-reviewers, editors, etc) has ever advanced a serious and substantive critique (https://2k4vbx44lajeo2rag2seu29o-wpengine.netdna-ssl.com/wp-content/uploads/2015/04/RNAi-Biosafety-DraftPaper-2015-LathamWilson.pdf).
But the other aspect is that, even if it becomes part of the published literature, it can still be ignored.
https://www.scientificamerican.com/article/how-chinas-bat-woman-hunted-down-viruses-from-sars-to-the-new-coronavirus1/
Notice
1) China says it has less infections and deaths now than it had in April
2) Shi says that the Mojiang miners suffered from a fungal infection when it was known almost immediately that they suffered from coronavirus.
3) Daszek is not willing to suspend his dangerous experiments despite the huge risks now evident to everyone.
4) Scientific American did not revise the numbers or comment on any of this when it republished the article. Nor has the scientific community subsequently.
Following these papers and discussion with great interest. Thank you for your courageous and intellectually honest efforts. One question that I did not see addressed: what would explain the highly stable nature of Covid-19 in humans? Does that suggest some sort of passaging process might have been undertaken on the original miner’s samples at WIV during the past few years, and that one of those more evolved/stable versions, rather than the raw miner samples, was what leaked?
If it’s from the lab samples of WIV as suggested, it’s naturally to think that the workers especially the ones in the Shi lab in the institute were the first to be exposed and possibly infected. Do you have any information or data regarding the number of infected cases or deaths within WIV? Any information regarding the possible pathway of virus traveling from the lab to the outside world?
The Economist this week (25 July edition) reports that Dr Daszak is suggesting another SE Asian country, such as Vietnam, Myanmar, Laos, to be a possible origin of the virus. From what you wrote, this might be viewed as diverting attention away from the Wuhan lab.
Perhaps you could share your last 2 papers with that publication?
It would be very interesting to trace, find and study the 3 surviving miners. This could be a huge breakthrough to support this hypothesis.
Excellent read. The one hole in the argument is passaging in the miners would be sufficient to produce SARS-CoV-2. Typical gof passaging in animals (e.g., ferrets) require at least 10 or more iterations. But the near perfect adaptedness in humans argues against passaging in animals like ferrets. The frightening implication is gof passaging experiments were conducted on HUMAN subjects. Given that prisoners are routinely murdered for their organs in China, the possibility of human passaging experiements cannot be ruled out.
We would say it is not a hole; rather, it is the part of the chain that is weakest in that it is least supported by evidence. On the other hand, the animal zoonotic origin thesis right now has effectively no support. In science you don’t go with a theory because it’s perfect you go with it because it is better than any other. No other theory comes close to meeting the basic requirements of the evidence accumulated so far.
Thanks you very much for this excellent detective work!
Please check, there seems to be something wrong here: “mutation at the 479 position in the SARS-CoV-2 RBD (corresponding to the 493 position in the SARS-CoV-2 RBD)”
Thanks for the spot! Fixed now.
May I ask if the link here is correct? I can’t seem to find any statement saying that the BSL-4 lab construction was delayed.
“Our supposition as to why there was a time lag between sample collection (in 2012/2013) and the COVID-19 outbreak is that the researchers were awaiting BSL-4 lab construction and certification, which was underway in 2013 but delayed until 2018.”
The link is what we meant to refer to. It says:
“The Chinese Academy of Sciences approved the construction of a BSL-4 laboratory in 2003, and the epidemic of SARS (severe acute respiratory syndrome) around the same time lent the project momentum. The lab was designed and constructed with French assistance as part of a 2004 cooperative agreement on the prevention and control of emerging infectious diseases. But the complexity of the project, China’s lack of experience, difficulty in maintaining funding and long government approval procedures meant that construction wasn’t finished until the end of 2014.”
And the lab seems to have opened in 2017.
Thank you for your research on this.
Thank you for this research.
Any thoughts on:
1. SARS-CoV-2 ability to remain infectious for days on surfaces (e.g . – plastic, stainless steel).
2. SARS-CoV-2 ability to remain infectious in aerosols?
3. are these traits of any other coronavirus?
4. are there genetic alterations which explain these traits?
5. if yes on 4, where did these alterations come from?
Could the surviving miners infected other individuals near the mine location? Is there any evidence of hospitalizations after these initial cases?
Another point: Have you attempted to convince the WHO to investigate this further? I was going to mention the CDC as well, but in this political climate the Chinese government would see it as furthering the Trump conspiracy theory.
I have one question. But first I want to thank the authors for their incredible scientific honesty and dedication to discovering the origin of this pandemic. The question is this, if the precursor to SARS-CoV-2 was able to adapt to humans so quickly in the lungs of miners so that there was little functional evolution when the virus was later unleashed on the world, then why didn’t SARS similarly wrap up most of its functional evolution in the first few people infected with SARS? SARS and MERS also infected the lungs and should have had many of the same forces accelerating their functional evolution that this work describes? At first, I thought that it could be explained by the diversity of viruses in mojiang cave and the assistance of recombination. Perhaps the viral inoculum that infected patient zero of SARS and MERS was only one virus or a less diverse viral population than the collection of viruses that infected each miner’s lungs at mojiang cave. I thought this was supported by some analyses that suggested that some regions of the SARS-CoV-2 genome were more similar to homologous regions of the genome recovered from pangolin samples than they were to RaTG13. So in other words I thought overall the SARS-CoV-2 genome was most similar to RaTG13 when considering its full length but when individual genes and regions are considered, some regions are more similar to the pangolin virus than to RaTG13. So I thought that there might be viruses in the mojiang cave that are more closely related to the pangolin virus than to RaTG13 despite being in the same cave and that these both infected the miners and recombined as they evolved to human hosts. However, this work suggests that RaTG13 is the sole progenitor of SARS-CoV-2. So what do you think? Why didn’t SARS and MERS evolve rapidly in the lungs of the first few people infected?
Thank you for the interesting article. I am puzzled by an apparent contradition, however. To quote your article, “ In our own search to resolve the COVID-19 origin question we chose to focus on the provenance of the coronavirus genome sequences BtCoV/4991 and RaTG13, since these are the most closely related sequences to SARS-CoV-2 (98.7% and 96.2% identical respectively). ”. Yet you also state both in the article and in a response to one of the early comments that those two strains are identical. How could they be identical to each other if they are not identically variant from SARS-CoV-2?
I would also like to suggest that similarity of the structures of two binding proteins does not necessarily suggest how close or far they are evolutionarily. Significant changes to the structured backbone and hydrophobic core of a protein are much more difficult than changes in loop or surface amino acid composition. I would have to know in more depth what you had in mind regarding the similarities between RBD structures.
Simple answer: BtCoV/4991 is identical to RaTG13 but not vice versa. This because BtCoV is a subset of RaTG13 in which region the homology is 100%
1.Efforts to uncover SARS-COV-2 origins ended;
2.Nobody is arguing over China’s responsibility on this tragedy;
3.Global cooperation against the disease ended;
4.Political war started using COVID-19 and how governments deal with it.
Brilliant hypothesis. I believe this is the best explanation for this mysterious disease. However, from a clinician’s viewpoint, it is interesting that with such a contagious and virulent viral illness why weren’t the medical personnel taking care of the 6 miners infected? For patients with infectious diseases one would use universal precautions but certainly not with the care we are taking with COVID-19. Also one could expect the families of the medical caregivers to also be exposed and sick. I’m thinking that the passaging that probably took place at the WIC led to an even more contagious disease. Any comments would be appreciated.
Here’s the problem. Because the CCP destroyed most of the early evidence and refused to allow “outside” virus experts into China to get at the origins of this virus was first exposed, they can make up anything now.
Of these, only patient 2 tested positive for Hepatitis and EBV.
Where these two patients Wu and Li?
“Their rhetoric is forthright but the evidence does not support that confidence.” I don’t want to be a jerk here but, this line could be said about your assumption that that WIV received this virus fully adept. To me its like you are just giving the WIV an out which is not unlike the Anderson study. If the WIV had it they gained it. That is what they do. Even in your study you conclude that level 2 labs can do this type of work. My understanding is that Reverse Genetics 2 is untraceable for signs of manipulation. So even from your convincing arguments that the furin site came from rapid evolution (Baric) it does not mean the lab did not manipulate this virus. There could easily be a statistical study conducted that shows how many hours worked on virus involves gain of function research.
I also think that you should have put in the fact that since you they talked to the Sars hero, whoever that is, that they took isolation very seriously and those elements that make this a highly infectious disease came after isolation. Otherwise if it was present upon infection it would have spread.
Outstanding and measured presentation FINALLY of a plausible explanation. Thank you. It also helps to logically explain why the CCP has been relentlessly trying to resist investigations into the origin. Dismissing the proximity of WIV as a coincidence in favour of the wet market distraction seemed illogical and a bit too unequivocal to my scientific background. I will do my small part to help spread the awareness of your thesis.
Dear Dr Latham and Dr Wilson,
Thank you for the very interesting article. After reading your article and the master’s thesis, may I make some comments.
No specific agent, fungal or viral, was found that caused the disease. Although the thesis’s conclusions strongly focus on the virus theory (understandable in view of SARS), details of individual treatments indicate that the doctors in charge treated the possibility of fungal infection at least equally seriously. All but one of the patients were treated with a combination of antiviral, antifungal and antibacterial medication. Strong possibility of fungal infection was suspected in at least two cases. Patient four in particular responded well to antifungal treatment. Patient five was specifically suspected of Histoplasmosis and responded well to antifungal treatment (“On May 6, according to the CT plain scan, the illness was getting worse. Therefore, we prescribed the antifungal medication and some hormone. The consolidation exudation in the upper lung has improved five days after. The temperature has dropped to normal. It indicates that antifungal med and hormones were effective” – Page 53.)
The only ground on which the thesis’ author rejected the fungal theory seems to be patient 6, (“The [only] patient [who] did not receive any anti-fungal medicine for treatment, yet still recovered. This suggested that the possibility of the illness being triggered by fungal infection is slim. – p. 57). However, this patient is 32 year old and has worked in the cave for only 4 days so had a good chance of recovery anyway.
On p. 62 the author again made a rushed conclusion: “The patient tested positive for Serum IgM by the WuHan Institute of Virology. It suggested the existence of virus infection”. However, IgM can also indicate histoplasmosis, a fungal infection (https://link.springer.com/chapter/10.1007/978-3-642-69943-6_21).
In conclusion, there is no information in the thesis that indicates that a virus is a more likely cause of the miners’ disease than a fungus.
So which one is more likely? The viral hypothesis requires that either MMP happened independently six times, or that it happened once and spread to other humans. MMP is viral evolution accellerated by hundreds of times, something that has never been observed in nature, so its likelihood is rather low. By the laws of probablity the chances of it happening six times independently is infinitesimal. Transmission from patient zero to the rest is therefore much more likely.
However, if human to human transmission was already possible and fast, as the cluster indicates, then why did the infection not spread further? No family member of medical personnel seemed to have caught the virus and it simply died out. Given what we know about Sars-Cov-2, it seems extremely unlikely. The thesis seems to imply that hospital staffs were quite unprepared for the situation and did not take any particular precaution (“Gaps and failings: […] The work of preventing hospital acquired infection should be the priority of ICU.” – p.62)
The fungal hypothesis seems to resolve these difficulties:
– Fungal infections can have the same symptoms as Covid-19 and often affect the lungs.
– Fungal pneumonia, which can be caused by several species, is often deadly among immunocompromised patients.
– Fungal infections often arise from bats and bird droppings, usually in caves. In fact, Histoplasmosis, a fungal infection, is often called the “cave disease”.
– Fungal infections are NOT CONTAGIOUS among humans.
– Fungal diseases is very common (at least compared to viral species jump), killing 1.5 million people every year (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5753159/)
On the basis of all these factors, I believe a fungal cause for the Mojiang miners (though not necessarily Histoplasmosis) is highly likely, and much more so than a viral one.
Reply to Quang Tuan Pham
The short answer to your question is that we did not claim that the miners certainly had coronavirus. There is some room for legitimate doubt.
The longer answer is that neither did we deny histoplasmosis and even it is in principle possible that the miners had both. They are not necessarily alternatives.
However, our main purpose in translating the Master’s thesis was to see, given that the title of the thesis mentions viruses only, how the author and colleagues reached those conclusions; i.e. were they reasonable? We think they were very reasonable but as I mentioned, it is still possible they were wrong. So while there is room for doubt, the overwhelming likelihood, as suggested by the subsequent queues of virologists at the Mojiang mine, is that it was a virus and likely a novel coronavirus.
On the second question you raise:
“MMP is viral evolution accellerated by hundreds of times, something that has never been observed in nature, so its likelihood is rather low. By the laws of probablity the chances of it happening six times independently is infinitesimal. ”
There seems to be a misunderstanding. Our thesis is not that every miner incubated SARS-CoV-2 (or something very similar), and thus relies on each evolving the virus, only that one of them did. However, it is possible (as you suggest) that someone in the mine developed the illness first and passed it to the others. In the article, we restricted our discussion to the most parsimonious possibilities, which places a lot of emphasis on evolution within the miners, but of course there are more complex versions, either prior to the mine outbreak or after the samples were taken to Wuhan, that are possible and indeed highly plausible.
I just was sent this 2 min clip: https://twitter.com/i/status/1287077979856572417
Zheng-li Shi saying (at last) that the “cave” where RaTG13 was collected by her lab was of interest due to “human cases”:
“we had a very great interest in this cave” and “this is a very, extremely, rare opportunity” which supports the idea that she suspected the cases were coronavirus.
Thank you for your efforts to alert the public to the dangers of risky coronavirus research (https://www.independentsciencenews.org/news/engineered-covid-19-infected-mouse-bites-researcher-amid-explosion-of-risky-coronavirus-research/) and for this fascinating work on the origins of SARS-CoV-2.
This paper and your previous one of June 2 pose some very interesting questions: questions, as you put it, not just about the “blame game,” but more importantly about the safety issues connected with some scientific research on viral pathogens.
However, I am curious about a different question: namely, would it make any difference to our ability to manage this epidemic if we knew more precisely who patient zero was or knew the original structure of the novel SARS-CoV-2 virus?
As you say in your June paper, “If the public has learned a lesson from the COVID-19 pandemic it is that science does not generate certainty. Do homemade face masks work? What is the death rate of COVID-19? How accurate are the tests? How many people have no symptoms?”
It is unsettling to me as a citizen and a parent to keep discovering how much we don’t know about the behavior of this virus which is doing so much damage to the country and to my family, but would it make any difference if we could, as you put it, unravel “the mystery of the origins of COVID-19”?
Best, Tom
Very interesting article. But I’m writing with grammatical objections. Incorrect use of the phrase, “beg the question” has spread throughout the English-speaking world like a virus. Where you use this phrase, you should have used “raise the question” instead. “Beg the question” refers to circular reasoning. https://www.txstate.edu/philosophy/resources/fallacy-definitions/Begging-the-Question.html#:~:text=The%20fallacy%20of%20begging%20the,called%20arguing%20in%20a%20circle.
is it conform with the assumed recombination history
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7217262/
https://www.biorxiv.org/content/10.1101/2020.04.20.052019v1.abstract
If the Mojiang miners were the source of Covid 19 then the Han Chinese have been exposed to it for many years. They should therefore be the most vulnerable to it and suffering the most infections and deaths. Instead the opposite is true….and you provide no explanation.
@Lifeguard Larry there is no explanation about other theories.
@yurieu I was unclear. My comment was directed at Jonathan Latham.
But – you’re right. His theory does not deal well with other objections … such as the one proposed above by Quang Tuan Pham.
We helped draft this letter asking for an international moratorium on gain-of-function research. We would really appreciate if scientists, lawyers and policy experts reading this article would consider signing on.
Please also share it with your relevant associates and acquaintances.
All shares much appreciated.
Jonathan
Link to the google doc:
https://docs.google.com/forms/d/e/1FAIpQLSdLCylWqArvdGSWf9ad42vG4euMikB4aJ5CEiWRD-bZOumfRQ/viewform
Have you addressed the fact that you didn’t see pandemic-like spread in the Kunming hospital or community from the mine workers? Do you think there was additional selective pressure in the Shi lab that led to a more virulent strain?
Allison answered this earlier:
“To answer Philip Ward: Good question. Our best guess is that it took a while for virus to evolve to be highly transmissible from human to human — this most likely happened later, after the miner(s) were in hospital. Also, once the miners were in the hospital the thesis is clear they were treated as if they had infectious diseases — perhaps even SARS or other very deadly diseases. Thus we expect the hospital staff were taking every precaution. There are other possibilities — the others were infected but had no symptoms or that infections were not made public or that we just have not encountered the data yet.”
And to your second question, the Shi lab presumably saw these samples as highly valuable. They would likely have studied and sequenced them but also we do not rule out enhancement at the WIV. Perhaps addition of the furin site?
I thank the authors for this article. The article complements the speculation about the RaTG13 virus of 2013 by Y. Deigin in the Medium article. I have a specific question in the third paragraph below. My background: I maintain a chain email of plausible “lab created”(aka ‘chimeric’) SARS-CoV-2 claims and will add this article to the list. I am a tech guy who has done patent work in biotech though it’s not my specialty.
I appreciate this article for the critique of the Kristen G. Andersen et al Nature Medicine March 2020 paper, which, prior to reading this, I’ve always maintained was polemical. I have speculated that biotech researchers wish to downplay the chimeric virus thesis since it might result in a moratorium, as happened in the USA around 2015 shortly on or before the UNC (Chapel Hill) SARS-CoV chimeric virus was designed by Menachery et al (including Shi and Baric). Analogous to law, you rarely see an article too critical of lawyers from most mainstream lawyers, probably for the same reason (professional curtesy and solidarity).
My specific question goes into creating a chimeric virus without traces, or signature-free (see, REF 1 below); why do this? This is referenced in REF 1 below which also favorably cites Latham and Wilson. My interest is that one thrust of the ‘SARS-CoV-2 is not lab-made’ argument is that the DNA sequence of the virus lacks the common signatures and traces found in recombinant DNA when said DNA is made from ligase and lyase agents such as one might buy from Bio Basic; i.e., SARS-CoV-2 ‘looks natural’ and not designed by a typical ‘gene jockey’. Since it’s been a number of years since I’ve dabbled in biotech, can somebody please comment on why biotech researchers even use signature-free and traceless clone technology? It’s almost like they are trying to cover their tracks? But perhaps there’s a innocent explanation, such as traceless and signature free DNA is more readily taken up by a host organism? Or is it just something that is done by biotech researchers because it can be done, for novelty’s sake? In short, somebody please explain why traceless, signature-free clone technology is employed.
-Ray Lopez
(REF 1) https://thebulletin.org/2020/06/did-the-sars-cov-2-virus-arise-from-a-bat-coronavirus-research-program-in-a-chinese-laboratory-very-possibly/#
“WIV’s first publication on the use of traceless, signature-free infectious-clone technology also appeared in 2016.32”
I am so glad to see the “Statement by Scientists…” Can non-professionals also sign? Is there a listing of the present signers available? Is the Cambridge Working Group (Consensus Statement 2014) involved?
On another topic, there was a comment by Ralph Baric (quoted in Yuri Deigin – April 22, 2020) that “Pangolins [strains of CoV2] have over 3000 nucleotide changes, no way they are the reservoir species. Absolutely no chance.”
Does this effect your argument in any way, assuming Dr. Baric retains his view?
Thank you for your very interesting article.
References:
April 22, 2020 – Lab-Made? SARS-CoV-2 Genealogy Through the Lens of Gain-of-Function Research – Yuri Deigin – Medium
https://medium.com/@yurideigin/lab-made-cov2-genealogy-through-the-lens-of-gain-of-function-research-f96dd7413748
TWiV 591: Coronavirus update with Ralph Baric
[3/13/20 – Ralph Baric – TWiV 591 audio – 26:50 – 28:25]
https://www.microbe.tv/twiv/twiv-591/
Excellent suggestion Milton. Here is the statement link:
https://docs.google.com/forms/d/e/1FAIpQLSdLCylWqArvdGSWf9ad42vG4euMikB4aJ5CEiWRD-bZOumfRQ/viewform
“Statement by Scientists, Lawyers, and Policy Experts on Why We Need a Global Moratorium on the Creation of Potential Pandemic Pathogens (PPPs) Through Gain-of-Function Experiments
Please fill out the form below to add your name to this letter……….”
(Btw, we helped draft this petition, which was floated in early Sept)
Recently Dr. Li-MENG Yan from Hong Kong published a paper hypothesizing a lab engineered origin of the virus. I wonder what you think of her analysis and how it might or might not fit with your ideas here. Thank you! https://zenodo.org/record/4028830#.X2AoSC2ZPoy
No Conspiracy
The experts have concluded
that c is from bats
and rodents or cats.
It certainly did not
drip from scientists’ vats.
The experts have concluded
only madmen and cranks
from crackers think tanks
could hatch a plot
of laboratory pranks.
The experts have concluded
three miners died
their doctors lied
in ’12 t’was mushrooms
not c bat pies.
The experts have concluded
that four-nine-nine-one
is no smoking gun
Shi sprang no leak
so Yan on the run.
The experts have concluded
Wuhan can be trusted
the prints have been dusted
no trace of a crime
no one need be busted.
Somewhere in the dark
conspiracies grow
they are hatched and nurtured
in cold neon glow
one lie is added and another and another
then leaked in the gutter
by nobody knows.
– MER
Hi,im no scientist,but according to sources in Wuhan(Personal)..people were getting sick with some unknown pneumonia type sickness in june/july 2019,which corresponds to people in the Usa getting sick with a unknown pneumonia type sickness according to Usa sources,which poses the question,did SARS-COV2 Infection start in the Usa To China..or from China To The Usa..in addition the disinfectant sprayed in Wuhan.by the Chinese Military is a disinfectant used exclusively to disinfect after a Bio Weapons Attack,so did the Chinese Military suspect there was a Bio Weapon Attack,in Wuhan?..or did they know?because Wuhan went into a complete Lock down,and elsewhere,the general consensus in Wuhan indicates that SARS-COV2 came from the Wuhan Lab Leak via a infected Lab Personnel,because within 7 days of the outbreak the Chinese military who deals with Bio Weapon Response took over all operations in Wuhan…there is lot of unknowns coming from china due to all info being blocked,incoming out coming,it points to a Chinese coverup,A-2-Z
Thank you! for your groundbreaking work and your bravery in pursuing the truth,
the really important science was always done only by people like yourself! not the conformist masses.
Good stuff here. Thank you for sharing. Any opinions on why male fatality is higher?
The miners were males and, according to the MMP theory, the virus evolved in one of them. Therefore it is specifically adapted to males. We decided to leave this suggestion out but it is an interesting observation. Thanks for the question.
Interesting article. And the perspective is also, especially the third rail of daring to suggest it could be that this was a lab escape event. When there have been thousands of them from the more than 200 labs housing pathogens and toxins. I do have a terminology question: what do you mean by ‘aerodigestive tract?’ about 3/4 way thru the article??
And btw it is all right to ‘beg the question’ – some woman commented on this being bad grammar or something so. I wonder if she ever heard of an ‘idiom?’ and you can still beg the question anyway. Nothing like rocks in the foundry.
Thanks for the question:
https://www.cancer.gov/publications/dictionaries/cancer-terms/def/aerodigestive-tract
so what do they ‘mine’ in this subject/suspect mine in Yunnan province?
copper, according to many sources though I haven’t assessed their credibility
This is the best thing I’ve read all year, and last year. It’s January 1st 2021 as I write this.
A few questions:
1. Do we know what year WIV started using their BSL-level 4 lab? The source in this article doesn’t seem to actually state what year WIV started using the BSL-4 facility.
2. Would it be plausible to infect non-human primates with high viral loads of RatG13 so that the evolution of the pathogens can be observed as a means of testing the miner passaging hypothesis? If it occurred in humans, shouldn’t we expect a similar outcome in closely related primate species?
Why were miners cleaning guano in a cave? Could it be they were hired to do so by WIV so that researchers could access the cave for bat samples with reduced risk of histoplasmosis? Might look into who those miners were hired by and who they worked for. If hired by WIV, there is motive to not report the illnesses of miners in the scientific reports. Next, those samples and tissues from the miners must be available somewhere. A PCR test on those samples would confirm the MMP hypothesis, would it not? Something is definitely fishy because Shi changed the name of the sample and seems to be intentionally not mentioning the miners.
Serum (and maybe other) samples were sent to the Chengdu Military disease centre, to Zhong Nanshan in Guangdong and to the WIV (Zheng-li Shi accepts they received 13 serum samples in her new addendum). All could be PCR’d or fully sequenced. Negatives would go a long way to refuting the thesis so it should be very surprising to anyone doubting this thesis that none have done this and published it long ago.
I just read the addendum. She says they tested the samples and did not test positive for sars like virus. But the phd thesis says otherwise, correct? She also says in the addendum they suspected viral infection of the miners. But another source she says she suspected fungal infection ????
Correct. And yes those are contradictory statements which cant easily be explained as changing her mind since there is no new data.
Is anything being done about this by WHO? Could testing the miners now reveal antibodies?
To my understanding nothing, they are only reviewing data given to them. What should be done is deep sequencing of those samples. If SARS-CoV-2 or an intermediate was present it would pretty much settle the matter. If coronaviruses were totally absent or just very different that would settle it in the other direction.
I’m not a virologist, a doctor, or in any way qualified to speculate on this issue but as the COVID-19 pandemic is by far the most globally disruptive event in my lifetime, I am quite interested in its origins. I am left with one burning question which is: If, as this article proposes, SARS-CoV-2 evolved from a bat coronavirus through some sort of accelerated natural passaging within the lungs of one or more of these miners; why did it not immediately break out by spreading to the healthcare workers who were treating them?
Andre that is exactly the question everybody asks and I don’t I don’t think the answer explains how the miners Getting there cool their COVID – like disease Adds up to being the origin of our pandemic. Anything is possible of course. But why didn’t more people get sick since 2013? Of course it is China and they will hide things but still I don’t think this is the answer.
1. Might COVID antibody testing of the Kunming Medical University personal that interfaced with the six ill miners back in 2012 answer Andre/Conrad question/comment?… or has too much time passed for test results to clearly answer these questions? Also, did the medical personnel utilize PPE from the get go while interfacing with the ill miners? (which as you allude to, review of the lab notebooks could be invaluable for answering these kinds of questions).
2. To shift gears forward, is it possible that “casual mask-wearing” has caused selection of these ‘new/latest’, more contagious, COVID variants? [where “casual mask-wearing” is defined/based on my own use of simple paper or cloth masks (which i’m thinking wouldn’t ‘cut it’ in the professional healthcare setting!) — just prior to going in to shop at a store, i pull the simple mask from my pocket and put it on… then as i walk out, i promptly remove it and insert it back into my pocket; on average, i’ll do this cycle with this one mask maybe 10 times… until i see the mask is too nasty to use anymore. So far, i believe i have not had the COVID; about three weeks ago, i had an antibody test done by the Red Cross as part of a whole blood unit donation; no one in our household has had symptoms; we’re waiting for an appointment to get in line for the COVID vaccination.]
sorry… too nebulous on item 2 in my 9:40pm, 21Jan2021 comment. my question is — is it possible that when a person has an initial uptake of COVID, and they continue to don/dof the simple mask… the virus mutates before the person begins to show symptoms; that is, before the person shows symptoms, the virus is now in/on their mask, and during the period between uptake and onset of symptoms, the virus has a medium/mechanism ideal for mutating; they exhale the virus onto the mask, it survives on the mask/medium, it is inhaled back into the lungs; during this cycling, a more contagious/harmful mutation is being incubated/’selected for’ during this cycling (need to recall the term you used for this type of mechanism in this, or your previous article [amplification?… something like that… i’ll look]).
Am I being too obvious if I say Prof Shi may have been deliberately vague while seeming to the point – she said
“Recently, we retested the samples …..against the SARS coronavirus 2 (SARS-CoV-2) ……and confirmed that these patients were not infected by SARS-CoV-2”
The wording does not say no other SARS – just no Covid-19
She then says
“We suspected that the patients had been infected by an unknown virus. Therefore, we and other groups sampled animals including bats, rats and musk shrews in or around the cave”
Again she does not say she did not find a match to the Miners unknown virus – she says what they found 9 beta coronaviruses but only identifies RaTG13 as sequenced
https://www.nature.com/articles/s41586-020-2951-z
You probably already have this file. For those who dont here is the link to Chinese copy of the Canping Phd.
Huang, Canping (2016) Novel Virus Discovery in Bat and the Exploration of Receptor of Bat Coronavirus HKU9.
https://drive.google.com/file/d/1fnoRmwl7d960UeOY2KqUG6e7TftqyXwP/view
Thanks! We are translating it. Has taken a bit of time due to the technical language.
Will your last presentation be on youtube or elsewhere soon ?
Talk III: The Pandemic Virus Industrial Complex 22nd Jan 2021
I thought the previous presentations were very interesting and clear
Thank you
Interesting article. Compared with tissue sample theory, how likely is “gain of function” research being the source of Covid? GOF was highlighted in recent publications, one of which was N.Wade’s.
Does anybody have the original MA thesis (outside of CNKI.net, don’t have access) to share, please? Thank you.
We did not post it online because the file is very large.
People who want it can email “info at independentsciencenews.org” or use the contact form
thanks!
Dear authors, your hypothesis has been mentioned and cited in this commentary article, recently published yesterday. Thank you for your excellent hypothesis.
https://www.frontiersin.org/articles/10.3389/fpubh.2021.702199/full?&utm_source=Email_to_authors_&utm_medium=Email&utm_content=T1_11.5e1_author&utm_campaign=Email_publication&field=&journalName=Frontiers_in_Public_Health&id=702199
The big flaws of this paper are the following:
1. No autopsy was performed on the miners who died in the Yunnan cave, and samples taken from their lungs prior to death did not isolate the virus or produce a sequence test that would have characterized the infection.
2.The recovered miners no longer carried the virus in their bodies and were not available to test for viral sequences, suggesting that the virus was not isolated from any of the patients from beginning to end, or that a new coronavirus or viral sequence was confirmed;
I hope main stream virologists will wake up from their dead sleep and do something with the findings in your excellent article!
The miners were treated with antivirals, which could increase the virus mutation rate, as with Remdesivir.
(Remdesivir was originally created and developed by Gilead Sciences in 2009, to treat hepatitis C and respiratory syncytial virus (RSV). – Wikipedia)
New article in support of yours:
“Compelling Evidence Suggesting the Codon Usage of SARS-CoV-2 Adapts to Human After the Split From RaTG13”
Yanping Zhang et al. Oct.2021
Thanks for pointing out the article. We had only just become aware of it. It does look promising and will definitely read.
RE: DT, “The big flaws of this paper are the following”, 3Oct2021 @ 4:33 PostMeridium
Welcome to China… most politically transparent nation on planet earth… good luck with that DT.